
不同林下植被覆盖度的马尾松种群结构与空间格局研究
2019-03-02 01:38孙眭涛王晓宁
福建农业科技 2019年12期
孙眭涛,王晓宁
(1.福建师范大学地理科学学院, 福建 福州 350007; 2.福建师范大学湿润亚热带山地生态国家重点实验室培育基地, 福建 福州 350007)
林下植被作为森林生态系统的重要组成部分之一,对森林群落演替、生态系统的稳定具有重要意义[1-2]。已有的研究发现,林下植被通过改变生境条件和资源竞争直接或间接地影响植物个体的存活、生长和繁殖更新,进而影响其种群动态。首先,林下植被的出现和发育通过调节林内微气候和改善土壤条件为乔木幼苗的发育提供适宜环境条件,同时林下植被的遮光效应也会限制底层幼苗对光的利用,进而阻碍种群的更新[3-6]。其次,林下植被与植株个体存在着资源竞争关系,虽然林下植被提高了土壤养分含量,但相较于乔木幼苗,林下草本植物具有更高效的养分吸收能力,这限制了乔木幼苗对养分的利用[7-10]。研究林下植被与乔木种群动态的关系,对于揭示变化生境条件下的种群动态机制具有重要的生态学意义。
种群结构是种群最基本的特征,是个体生存能力与其生存环境长期适应的结果,它不仅可以反映种群内部的个体大小分布情况,也能分析种群的受干扰状态以及预测未来种群动态和演替趋势[11]。种群的空间格局受物种自身的生物学特性、种内种间关系、生境条件以及其相互作用的影响,是种群的基本数量特征之一[12-13]。研究森林群落中优势物种的种群结构与空间分布格局不仅可以阐明种群的动态变化和格局的成因,也可为当地的生态恢复和植被恢复提供一定的理论依据。
马尾松Pinusmassoniana是我国亚热带红壤水土流失区生态恢复与重建的乡土树种和先锋树种,其动态变化指示着亚热带森林群落的演替阶段和恢复水平[14-15]。自20世纪80年代以来,南方地区实施了以重建地表植被,缓解水土流失为目标的大规模植被重建工程。经过30多年的水土流失治理,该地区逐步形成了以马尾松为主的纯林和混交林,其中以福建省长汀县最为典型[16]。随着生态恢复水平的提高,林下植被的出现和发育必然会对马尾松种群产生不同程度的影响。本研究以福建省长汀县的马尾松种群为研究对象,研究其在不同林下植被覆盖度的种群结构与空间分布格局,以期为亚热带水土流失区马尾松林的管理与改造提供一定的科学依据。
1 材料与方法
1.1 研究区概况
研究地位于福建省西南部长汀县河田镇(116°16′~116°30′E,25°18′~26°02′N)。属中亚热带季风性气候,年平均温度20.5℃,年平均降水量1 700 mm,年平均降雨日数166.5 d,降雨主要集中在4月至6月,该时期的降水量占全年降水量的60%以上,7月至8月受副热带高气压带的控制,降水稀少且不稳定,容易形成短期的干旱或洪涝。地貌以低山、丘陵为主,地形起伏大[17]。土壤类型为花岗岩风化发育的山地丘陵红壤土。地带性植被为季风气候下发育的亚热带常绿阔叶林。由于历史上的大规模植被破坏,该地区形成了中国南方最典型的红壤水土流失区。经过30多年的生态恢复,现存植被主要为以马尾松为主的纯林和针阔混交林[18]。
1.2 样地设置与野外调查
为了尽可能找到除林下植被覆盖度指标外生境相似的样地,2018年7月在福建省长汀县河田镇设置了3个900 m2的马尾松种群监测样地,分别为Luvc(低林下植被覆盖度样地)、Muvc(中林下植被覆盖度样地)、Huvc(高林下植被覆盖度样地),每个样地的林分相似,马尾松密度、株数接近,且样地间距不超过2 km。按照“相邻网格法”将样地划分为5 m×5 m的小样方。测量每一个小样方内的马尾松个体并挂牌标记,树高大于1.3 m的记录胸径(mm)和基径(mm),树高低于1.3 m的记录其树高(cm)和基径(mm)。统计所有马尾松个体的松果数和坐标位置。记录每个小样方的林下植被覆盖度信息、生物多样性等生境信息。样地基本情况见表1。
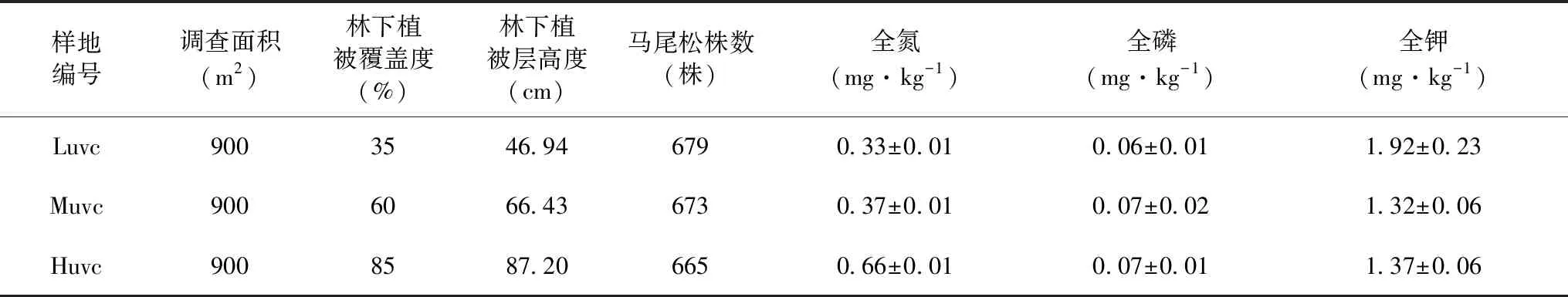
表1 马尾松样地基本信息情况
1.3 调查方法
1.3.1种群结构动态与趋势预测 正常条件下,马尾松基径大小与其树龄成正比。本研究采用空间推时间,径级结构代替年龄结构的方法进行种群结构分析。根据马尾松基径(BD)的调查数据,除第Ⅰ、Ⅱ龄级外,按30 mm为间距,将基径划分为12个等级:BD≤5 mm为I级;5 mm 依据马尾松的生活史特点,将其划分为4个生长阶段:幼苗(Ⅰ级)、幼树阶段(Ⅱ~Ⅲ级)、成树阶段(Ⅳ~Ⅶ级)、大树阶段(Ⅷ~Ⅻ级)。 1.3.2种群空间分布格局的确定 种群的空间分布格局是植物物种本身与环境相互作用最直接的反应。本研究通过计算7个指标来判定不同林下植被覆盖度的种群空间格局类型和聚集强度[19-20]。 (1)负二项参数(K) K值代表聚集程度,v为方差,m为均值。K值越小,表示种群聚集的程度越大;当K小于0时,种群为均匀分布。 (2)丛生指数(It) It<0为均匀分布;It=0为随机分布;It>0为聚集分布。 (3)平均拥挤度(m*) m*值表示样方内部植株个体所受的拥挤程度,m*值越大则聚集强度越大,植株个体受其他植株的拥挤效应也就越大。 (4)聚块性指数(m*/m) m*/m<1为均匀分布,m*/m=1为随机分布,m*/m>1为聚集分布。 (5)Cassie指数(Ca) Ca=1/k Ca<0为均匀分布,Ca=0为随机分布,Ca>0为聚集分布。 (6)扩散系数(Cd) 扩散系数为检验种群是否属于随机型的一个系数,当Cd<1时为均匀分布;Cd=1为随机分布;Cd>11为集群分布。 (7)Morisita指数(Iδ) 如果Iδ<1,则趋于均匀分布;Iδ=1,为随机分布;Iδ>1,则趋于集群分布。 利用Excel 2007和SPSS 19.0软件对数据进行统计分析。采用Excel 2007和Origin 9软件统计制图。 由图1可知,3个不同林下植被覆盖度的马尾松种群结构差异明显,各样地马尾松种群年龄结构的特征具体为: 低林下植被覆盖度(Luvc)样地的马尾松种群结构不完整,表现为:幼苗(Ⅰ级)数量最多,个体数占总数的61%以上;幼树(Ⅱ~Ⅲ级)数量次之,占总数的21%;成树(Ⅳ~Ⅵ级)数量少,占总数的17%;大树(Ⅶ~Ⅻ级)个体缺乏。年龄结构成基部极宽顶部狭窄的金字塔型,增长性强,为增长种群。由于幼苗和幼树的数量多,未来马尾松种群扩张速度很快。 图1 不同林下植被覆盖度的马尾松种群龄级结构Fig.1 Age structure of Pinus massoniana population under different understory vegetation coverage注:a为低林下植被覆盖度(Luvc);b为中林下植被覆盖度(Muvc);c为高林下植被覆盖度(Huvc)。 中林下植被覆盖度(Muvc)样地的马尾松种群结构最为完整,表现为:幼苗(Ⅰ级)植株数量最多,占种群的68%以上;幼树(Ⅱ~Ⅲ级)的数量次之,占种树的21%,成树和大树个体(Ⅳ~Ⅻ龄)的分布较为均匀,个体数量变化不大;年龄结构与低林下植被覆盖度(Luvc)样地的年龄结构相似,同样成基部宽顶部窄的金字塔型,为增长种群。 高林下植被覆盖(Huvc)样地的马尾松种群结构较为完整,表现为:幼苗(Ⅰ级)数量少,仅有5株;幼树(Ⅱ~Ⅲ级)数量最多,占总数的75%以上;成树(Ⅳ~Ⅵ级)数量较多,占总数的16%以上;大树(Ⅶ~Ⅻ级)数量甚少,仅占总数的3%。高林下植被覆盖度(Huvc)样地的马尾松种群大体为纺锤形,幼树和成树占有相当大的比例,绝大多数个体的生理特性已经成熟,在一定时期内能够继续生长繁殖,但由于马尾松幼苗的缺失,因此,种群已经表现出衰退的征兆。未来高林下植被覆盖度(Huvc)样地的马尾松种群衰退演替的速度会加快。 马尾松种群存活曲线(Sc)表明:不同林下植被覆盖度的马尾松种群存活存在较大差异。具体表现:低林下植被覆盖度 (Luvc) 样地和中林下植被覆盖度(Muvc)样地的马尾松幼苗、幼树的存活率较高,随着径级的增大,存活概率逐渐下降(图2)。马尾松前期个体较小,对空间资源竞争较弱,随着径级的增大,个体的资源需求量增大、自疏效应增强及环境筛的强度增大,导致存活率下降,存活个体减少。高林下植被覆盖度(Huvc)样地的马尾松种群存活曲线趋于Deevey-Ⅰ型,表示个体成熟前只有少量个体死亡。幼苗-幼树的存活概率上升,幼树-成树的存活概率不断下降,当径级达到Ⅸ级时,存活概率上升。 植物种群分布格局是对植物自身的生物学特性、种间关系以及生存环境等因素相互作用下的种群个体水平空间配置和分布状态的定量描述。 根据表2结果,不同林下植被覆盖度的马尾松种群空间格局类型均表现出了集群分布的特征。但聚集强度在不同样地存在显著差异。具体表现,3个样地中方差/均值(v/m)、聚块性指数和Morisita指数均>1,平均拥挤度均大于各自的平均值,表明不同林下植被覆盖度的马尾松种群分布类型为典型的集群分布格局。从不同样地的聚集强度分析,低林下植被覆盖度样地的种群聚集强度要大于其他两个样地,表现为低林下植被样地(Luvc)的m*值显著大于中林下植被覆盖样地(Muvc)和高林下植被覆盖样地(Huvc)。 表2 不同林下植被覆盖度下的马尾松种群空间分布格局分析 注:类型“C”为聚集分布(Clumped distribution)。 种群的年龄结构和空间分布格局是对外部环境适应性的体现,是特定条件下种群长期发展的结果。不同林下植被覆盖度下的马尾松种群结构呈现出不同的种群结构特点。低林下植被覆盖度和中林下植被覆盖度的样地上马尾松种群属于增长型,高林下植被覆盖度样地的马尾松种群为衰退型。在低林下植被覆盖度的样地中,马尾松种群呈现出典型的金字塔型径级结构。野外调查发现,在林下植被较少的地区,容易形成连片的裸地,马尾松喜光照、耐贫瘠干旱的先锋植物特性使其容易在裸地上生存,马尾松幼苗大量萌发,但地表的裸露使马尾松幼苗面临极端的水热胁迫,尤其是7月至8月份高温少雨的天气条件,地表干热化程度严重,加上幼苗根系浅,不成熟,抵抗极端环境的能力差,只有约40%的新生幼苗可以成长至幼树阶段,这说明稀少的林下植被虽然可以使马尾松获得充足的光照资源,但也使其容易面临极端的干热化胁迫[15]。随着林下植被的出现和发育,马尾松幼苗数量出现轻微减少,可能是由于马尾松是一种强阳性植物,上层乔木和林下植被的遮阴作用使部分幼苗出现死亡,成树个体的数量显著提升,表明林下植被一定程度上改善了马尾松的生存环境。然而在高林下植被覆盖地区,低于林下植被高度的马尾松个体无法获得充足的光照,以及激烈的种内种间资源竞争,最终导致小径级的马尾松在种群结构中比重下降。小径级马尾松大量死亡,这表明种群已步入衰退状态,可以预见,马尾松种群会随着演替进程的推进逐渐被其他阔叶树种所替代[21]。 马尾松种群的空间分布格局主要是由于自身的生物学特性和其生存环境共同决定[22]。3个样地的马尾松种群均呈集群分布,但聚集强度随着林下植被覆盖度的提高有所降低,这与其生存环境不同有关。在低林下植被覆盖地区,群落物种组成单一,主要以马尾松为主,马尾松幼苗多分布于母树的周围,这主要是由于马尾松松果多散落于大树周围,这种高强度的聚集分布也有利于小径级个体的存活和生长。随着林下植被覆盖度的提升,如芒萁、木荷、胡枝子等植物逐步进入群落内部,使得种内和种间竞争加剧,特别是对光资源的竞争,低于林下植被层高度的马尾松个体出现死亡。因此,到了高林下植被覆盖阶段,马尾松种群的集群强度出现下降,主要是由于部分小径级个体的死亡所导致。不同林下植被覆盖阶段的马尾松种群空间格局变化反映了马尾松在生长发育过程中的一种生态策略和适应机制[23]。 植物与其生存环境是一种相互影响相互依赖的关系[24],马尾松种群会不断地调整其生存策略以适应生态恢复过程中的变化生境。马尾松属耐旱、耐贫瘠生境的阳性树种,若地表植被覆盖量少,极端的地表干热化胁迫接近甚至超过了马尾松幼苗的耐受性,使其光合速率下降、生长放缓甚至枯萎死亡;若林下植被覆盖过多,致密的林下植被层使阳光难以抵达植被底层,光资源则成为马尾松种群增长的主要限制因子。1.4 统计分析
2 结果与分析
2.1 不同林下植被覆盖度下的马尾松种群结构、存活曲线分析

2.2 不同林下植被覆盖度的马尾松种群空间格局分析

3 讨论与结论
猜你喜欢
科学技术创新(2022年30期)2022-10-21
农业与技术(2021年23期)2021-12-14
黑龙江水利科技(2020年8期)2021-01-21
原子与分子物理学报(2020年5期)2020-03-17
河北果树(2020年1期)2020-02-09
绿色科技(2019年5期)2019-11-29
现代园艺(2017年11期)2017-06-28
广西林业科学(2016年1期)2016-03-20
广西林业科学(2016年1期)2016-03-20
广西林业科学(2016年1期)2016-03-20
