
一株鸽新城疫病毒Pi/YN/CH/0122/2018株的分离鉴定
2019-02-22 08:48李敏和春林严红亚信爱国常志顺
云南畜牧兽医 2019年1期
李敏,和春林,严红亚,信爱国,常志顺*
(1. 云南省畜牧兽医科学院 养禽与禽病研究所,云南 昆明 650224;2. 砚山县维摩乡畜牧兽医工作站,云南 砚山 663100;3. 丽江市古城区大东乡农业综合服务中心,云南 丽江 674100)
新城疫是由新城疫病毒(Newcastle disease virus,NDV)感染引起的一种急性、烈性、高度接触传染性禽类传染病,被世界动物卫生组织(OIE)列为必报传染病之一[1]。NDV属于副黏病毒科,是一种单股、负链、线性、不分节段的RNA病毒,只有一个血清型,但因宿主范围、毒力不断演变,NDV 给养殖业带来了巨大的经济损失。NDV根据致病力差异,可分为弱毒株、中等毒力株和强毒力毒株三种类型。
鸽新城疫,最早被称为鸽副黏病毒病,是由I型禽副黏病毒引起的一种高度接触性、败血性传染病。该病病原与鸡NDV在血清学上没有显著差异,两者基因序列同源性较高。随着时间的推移及饲养方式的改变,鸽Ⅰ型副黏病毒呈现流行广、传播快的特点。迄今为止,我国华南、华北、华东等地区大部分省份都有该病的报道,给我国养鸽业造成了重大的经济损失[2]。
本实验通过对云南某赛鸽公棚疑似自然感染NDV的病死鸽采集的病料进行病原分离鉴定及分子流行病学分析,旨在为临床诊断NDV感染提供实验室检测依据,了解云南省鸽源新城疫病毒感染情况以及为防控提供理论依据。
1 材料与方法
1.1 病料
2018年5月,云南某赛鸽公棚出现幼鸽发病,发病率12%,死亡率8%,送到本研究所进行临床解剖及实验室诊断,采集病料进行分离鉴定。
1.1.1鸡胚和NDV病毒
9日龄非免疫健康鸡胚本研究所提供,LaSota株疫苗毒,本研究所保存。
1.1.2阳性血清
NDV、禽流感病毒(H5、H7、H9亚型)标准阳性HI血清购自中国农业科学院哈尔滨兽医研究所,减蛋综合征病毒(EDSV-76)HI血清由本研究所制备保存。
1.2 分子生物学试剂及引物合成
RNA提取试剂盒购自天根生化科技(北京)有限公司。PCR所用的分子生物学试剂均购自TaKaRa公司。参考新城疫诊断国家标准(GB/ T 16550-2008)合成引物[3],扩增上游引物:5′-ATGGGCYCCAGAYCTTCTAC-3′,下 游 引 物:5′-CTGCCACTGCTAGTTGTGATAATCC-3′。引物由上海生工生物工程技术服务有限公司合成。扩增产物直接送昆明硕擎生物科技有限公司测序。
1.3 病毒分离
采取病死鸽的肝、肺脏组织,剪碎,加灭菌生理盐水研磨,反复冻融3次,加入双抗(青霉素和链霉素)1000IU/mL, 4℃过夜,12000r/min离心5min,取上清液经尿囊腔接种9日龄非免疫健康鸡胚,置37℃孵化器内培养7d,每12h观察一次鸡胚死亡情况。24h后死亡以及结束孵化时存活的鸡胚置4℃冰箱4~24h,收获尿囊液备用,观察胚体病理变化。
1.4 病毒鉴定
1.4.1血凝(HA)和血凝抑制(HI)试验
按照常规方法[4]测定收集的尿囊液的HA效价,并以抗NDV、EDSV、H5、H7、H9标准阳性血清进行HI试验。
1.4.2病毒MDT测定
参考国家标准(GB/ T 16550-2008)进行 MDT 测定[3],将病毒尿囊液用灭菌生理盐水连续10倍稀释;每个稀释度经尿囊腔接种5枚9日龄的SPF鸡胚,每枚按0.1mL/接种,置37℃孵化器内培养,弃去24h内死亡胚,以后每6h照蛋一次,连续观察7d,记录各鸡胚的死亡时间。按Karbre氏法计算EID50,并根据所有胚胎死亡的最高稀释度计算MDT。
1.4.3病毒RNA的提取及PCR扩增
取收集的病毒鸡胚尿囊液300μL于灭菌的3mL EP管中,再加入1000μL Trizol试剂,混匀,室温静置10min;加入200μL氯仿,轻轻混匀,室温作用10min;4℃、1200r/min离心15min;取上清,加入上清相同体积的异丙醇,振荡混匀后室温静置10min;4℃、1200r/min离心10min;弃上清液,加 入1000μL 75%乙醇洗涤;4℃、1200r/min离心5min,弃去上清液,室温静置晾干,加40μL ddH2O溶解。提取的RNA立即进行RT-PCR,RT-PCR步骤按试剂盒说明书进行,PCR反应条件为94℃ 3min,94℃ 30s,55℃ 30s,72℃ 45s,进行35个循环,最后再72℃延伸7min。取5μL反应产物进行2%的琼脂糖凝胶电泳成像观察。
1.4.4 PCR产物核酸序列测定及进化树的构建
按DNA胶回收试剂盒的操作进行PCR产物回收,送公司测序,参考已报道的方法[5],根据已发表的NDV主要参考毒株构建该毒株的系统进化发育树,所用NDV参考毒株信息见表1。
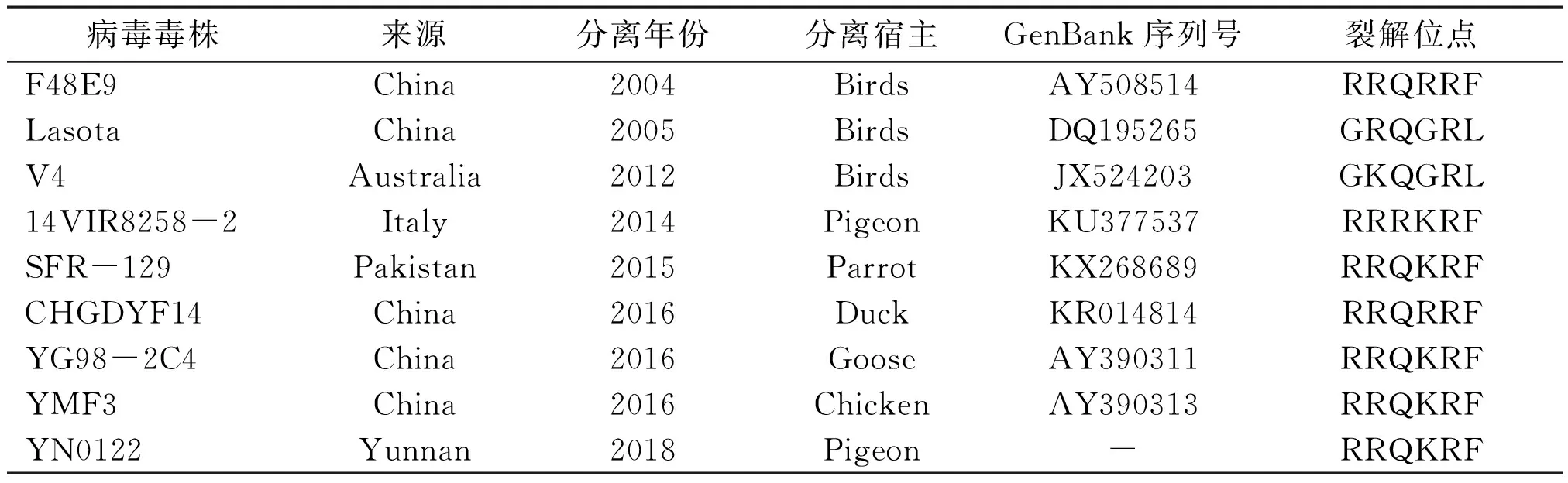
表1 所用NDV参考毒株信息
2 结果与分析
2.1 发病鸽临床症状及剖检情况
经临床解剖及流行病学调查,初步诊断为疑似新城疫感染。发病鸽临床症状表现为精神萎靡,羽毛蓬乱,体温升高,食欲下降,咳嗽,呼吸困难,有黏性鼻液,张口呼吸,拉黄色或绿色稀便,翅膀下垂,有扭头、转圈等神经症状,后期病鸽因瘫痪无法采食,衰竭死亡。耐过鸽扭头、转圈等神经症状明显。剖检可见全身黏膜出血,淋巴结肿大、出血,腺胃黏膜出血、坏死,盲肠扁桃体肿大、出血,脑组织充血水肿。该公棚鸽发病前1周进行过颈部皮下0.5mL/羽LaSota 株灭活疫苗免疫,经检测,发病鸽血清NDV抗体HI效价为 24~26。
2.2 接种后死亡胚体的病变观察
接种24 h后鸡胚开始陆续出现死亡,鸡胚死亡时间为接种后24~96h,胚体全身性出血、水肿,头颈部、足趾和翅膀出血严重,见图1。
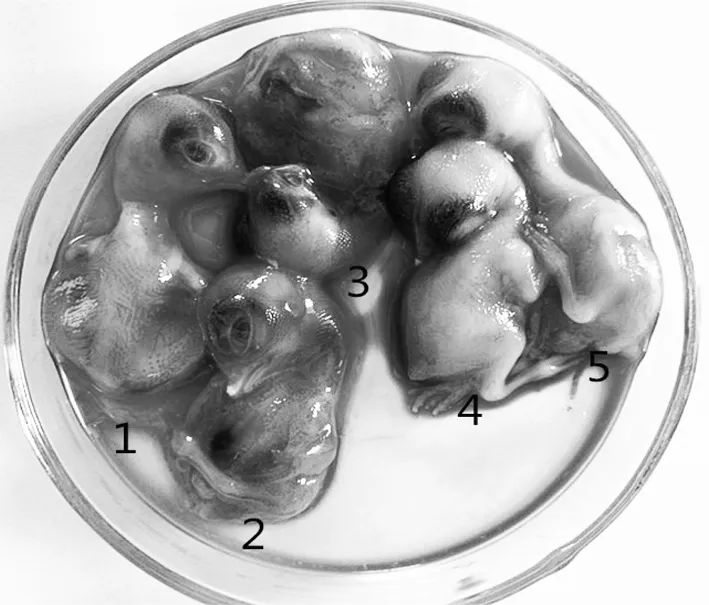
1、2、3为感染病毒后鸡胚,4、5为正常鸡胚
2.3 病毒鉴定
鸡胚尿囊液毒可凝集鸡红细胞,鸡胚死亡时间为接种后24~96h,其血凝性可被 NDV阳性血清抑制,不能被EDSV、H5、H7和 H9亚型的阳性血清抑制,证明所分离到的病毒为新城疫病毒,命名为Pi/YN/CH/0122/2018(简称YN0122)。
2.4 EID50及 MDT 的测定结果
盲传2代后尿囊液毒HA效价为7log2,对鸡胚的 EID50为 10-7.5/ 0.1mL;MDT为 66h,最小致死量为10-6,结果表明该分离株为中等偏强毒力毒株。
2.5 RT-PCR扩增结果
对提取的病毒尿囊液RNA进行PCR扩增,经2%琼脂糖凝胶电泳观察,结果得到约500bp目的核酸片段,与预期结果相符(见图2)。
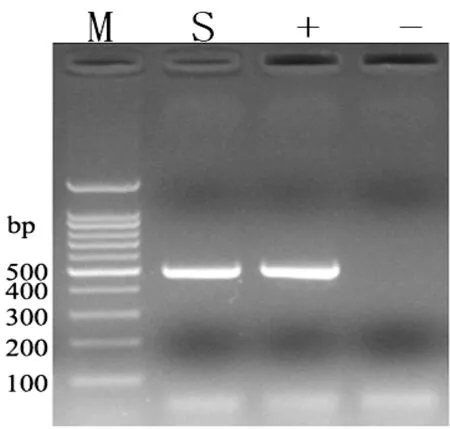
M:DNAMark;S:分离株病毒;+:NDV LaSota 株阳性对照;-:阴性结果对照
图2目的片段PCR扩增结果
2.6 核酸序列比对分析
测定上述PCR扩增产物核酸序列,应用DNAStar 6.0对YN0122进行序列拼接与比对分析,并用MegAlign软件构建系统进化树(见图3)。结果表明,分离株YN0122与传统NDV疫苗LaSota株和标准强毒株北京株F48E9、NDV V4株、鹅副黏病毒病代表株YG98-2C4的核苷酸序列同源性分别为81.2%、82.4%、81.4%和87.2%,同源性较低,遗传距离较远,而与鸽源Italy分离株14VIR8258-2同源性为91.3%,遗传距离较近。该毒株F基因裂解位点氨基酸序列为112R-R-Q-K-R-F117,与NDV强毒株的F 基因裂解位点的氨基酸结构一致,见表1、表2。
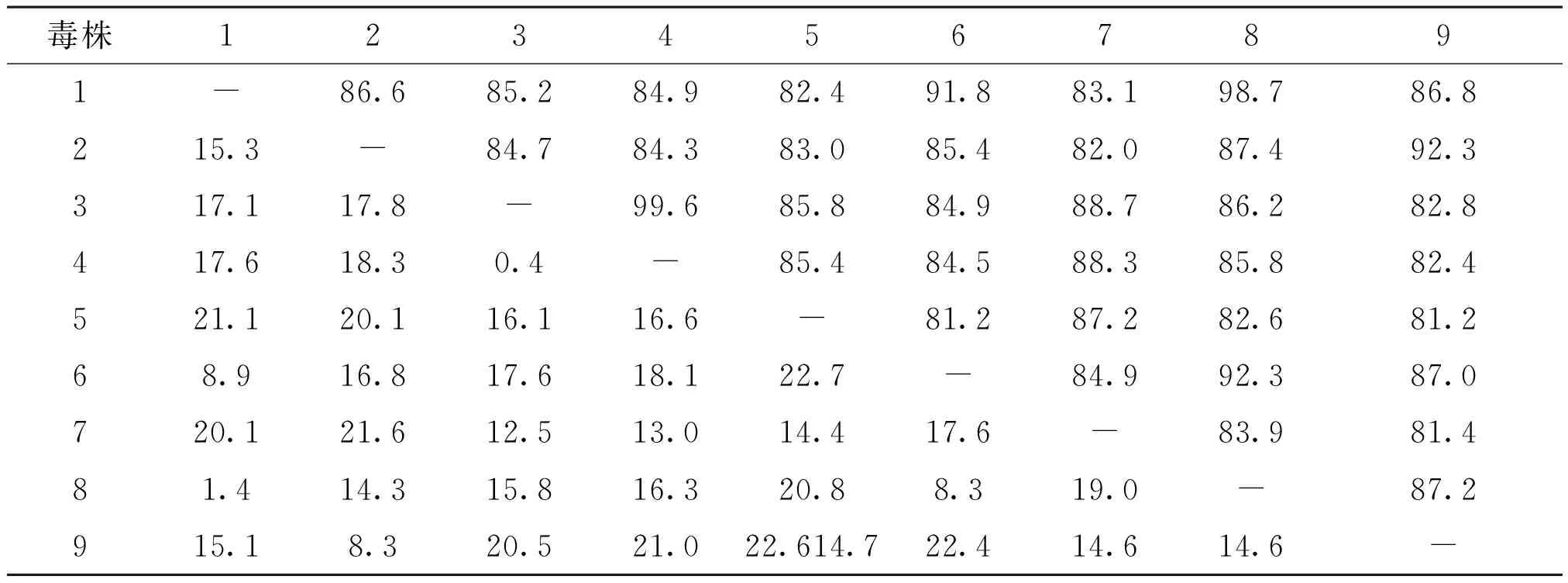
表2 9株 NDV F基因核苷酸同源性的比较
1:YMF3(AY390313);2: 14VIR8258-2(KU377537);3: CHGDYF14(KR014814);4:F48E9 (AY508514);5:Lasota(DQ195265);6: SFR-129(KX268689);7: V4(JX524203);8: YG98-2C4(AY390311);9: YN0122
图3基于NDV F基因的遗传进化树
3 小结
本试验采用传统的鸡胚接种方法分离病毒,病料接种鸡胚盲传 2 代后,发现收获的鸡胚尿囊液具有血凝活性,能与红细胞发生凝集反应,其 HA 效价为 24~26。HI 试验结果证明,该分离株的血凝活性只能被NDV 标准阳性血清所抑制,初步鉴定该分离株为NDV。F2代尿囊液毒EID50为 10-7.5/ 0.1mL;MDT为 66h,最小致死量为10-6。用RT-PCR检测,得到预期特异性扩增目的片段。明确了从病死鸽子中分离到的是一株NDV。根据扩增片段核酸系列比对分析证实,分离株YN0122 F基因编码的氨基酸裂解位点序列为112R-R-Q-K-R-F117,与NDV强毒株的F基因裂解位点的氨基酸结构一致,YN0122株具有中等毒力偏强的特征。
鸽新城疫病毒又称鸽Ⅰ型副黏病毒,属于副黏病毒科、副黏病毒属,只有一个血清型,但不同毒株的毒力表现出很大的差异[5],鸽Ⅰ型副黏病毒和鸡新城疫病毒同为一属,含有极为相似的抗原成分[6]。本实验所分离到的鸽新城疫毒Pi/YN/CH/0122/2018株,根据试验结果初步判定为中等偏强新城疫病毒,但有文献报导,鸽新城疫病毒(鸽Ⅰ型副黏病毒)的毒力和致病性不能完全由F基因裂解位点的特征性结构决定[7],主要通过动物试验的方法进行测定[8]。因此,要弄清楚分离株的毒力和致病性,还需进行动物回归试验。
此次发生疫情的赛鸽公棚免疫过LaSota 株灭活疫苗,正常及发病鸽新城疫血清抗体HI效价达 24~26。针对本次鸽源NDV感染不能提供完全的保护,可见,尽管NDV只有一个血清型,但不同毒株的毒力存在差异;本试验YN0122株F蛋白基因与LaSota株同源性为81.2%,同源性较低,遗传距离较远,生产实践中使用LaSota疫苗对鸽进行免疫存在免疫失败的风险。因此,了解鸽NDV的生物学特性,为新城疫的防制提供重要的理论依据。
鸽新城疫是一种急性、烈性传染病,给赛鸽公棚造成了严重的经济损失。赛鸽公棚每年春季会从全国各地收购幼鸽,集中饲养、训练后参加秋季的赛飞比赛。复杂的幼鸽来源背景、疫苗免疫情况以及赛鸽在野外飞行训练过程中,易接触到携带有新城疫病毒的野生鸟类,此养殖模式易使新城疫等传染病发生并在鸽群中扩散。因此,研究制定并实施包括新城疫在内的疫病综合防控措施对公棚赛鸽养殖尤显重要。
猜你喜欢
农药科学与管理(2019年9期)2019-11-23
农药科学与管理(2019年6期)2019-11-23
家禽科学(2019年2期)2019-06-11
食品与生活(2018年7期)2018-09-19
中国畜禽种业(2018年10期)2018-01-19
中国介入影像与治疗学(2017年12期)2017-01-14
中国畜牧兽医文摘(2015年9期)2015-12-29
现代泌尿外科杂志(2015年9期)2015-02-20
特产研究(2014年4期)2014-04-10
中国兽药杂志(2013年11期)2013-11-23
