
滇西北高原湿地景观变化与人为、自然因子的相关性
2019-02-20 10:38:00王泉泉张卫国王志保肖德荣
生态学报 2019年2期
王泉泉,王 行,张卫国,王志保,肖德荣
西南林业大学国家高原湿地研究中心/湿地学院, 昆明 650224
自20世纪50年代以来,由于自然因素和人为活动干扰,全球湿地面积急剧减少[1]。我国第二次全国湿地资源调查结果显示,在2003—2013年10年间,我国湿地面积减少了339.63万hm2,减少率达8.82%[2]。研究发现,以气候变化为主导的自然因素、以社会经济发展为表征的人为活动干扰,是导致湿地分布面积不断减少的主要驱动因素[3-4]。研究自然因子变化与人为活动干扰在湿地生态系统演化过程中的作用与贡献,是掌握两大驱动因子对湿地生态系统的作用途径与机制的基础。
高原湿地为我国“两屏三带”生态安全格局的重要组成,在水源涵养、生物多样性保护和碳固定等方面发挥着不可替代的作用[5]。但是,高原湿地生态系统极为脆弱,易受外界的干扰而导致湿地结构改变和生态功能退化[6]。近几十年来,以气温升高和降水减少为主要特征的全球气候变化,导致高原湖泊水位下降、面积减少[7],雅鲁藏布江径流减少[8],沼泽湿地萎缩退化[9],一些区域沼泽湿地停止发育,部分地段沼泽泥炭地出现干燥裸露的现象,甚至沙化和荒漠化[10]。同时,过度放牧、排水垦殖、泥炭开采以及湿地旅游资源的无序开发等[11- 12],进一步加剧了高原湿地生态系统破坏,致使高原沼泽湿地发生逆向演替[11]。因此,气候变化与人为活动干扰,已导致我国高原湿地面积分布总量下降、湿地生境改变、功能退化,严重威胁着湿地区域人类生存环境和生物多样性保护[13- 15]。
滇西北高原湿地地处青藏高原东南缘,是我国“川滇生态屏障”的重要组成部分,对气候变化高度敏感[16]。自1960—2011年,滇西北高原地区大气年均气温上升了1.3 ℃,气温上升率达22.9%,其气候变暖的总体趋势与全球变化趋势一致,但显著高于云南其他地区[17]。另外,随着该区域社会经济的快速发展,人为经济活动对湿地生态系统的干扰类型不断增多、干扰强度也不断加大。其中,过度放牧、排水围垦、湿地旅游资源的无序开发等人为干扰活动,直接导致湿地分布面积不断萎缩,湿地土壤理化性质和养分,以及植物组成、类型等发生改变[18-20];汇水区森林植被的破坏,致使湿地区泥沙淤积、水文改变[21]。气候变化与人为经济活动的干扰,威胁着该区域湿地生物多样性的保护。
本研究选取滇西北高原湿地纳帕海为研究对象,基于面向对象的分割技术、结合目视解译和实地验证方法,并收集当地28年(1987—2015年)来的气温、降水等自然因子,以及社会经济发展指标等人为活动干扰因子,研究纳帕海汇水区(1)湿地类型及分布,(2)景观多样性,(3)湿地分布面积与景观多样性变化和气候因子、社会经济发展指标的相互关系及解释度。通过研究,拟掌握滇西北高原纳帕海湿地分布及景观变化的自然与人为贡献率,为揭示两大驱动因子对湿地生态系统的作用机制奠定基础。
1 材料与方法
1.1 研究区域概况
纳帕海位于滇西北横断山脉中段香格里拉市境内(99°37′—99°41′E,27°49′—27°55′N),最低海拔为3260 m(图1)。该区域属寒温带高原季风气候区,受青藏高原寒流影响,干湿季分明。其中,6—10月份为雨季,11月—翌年5月为干季。年均降雨量619.9 mm,年均温为5.4℃,最冷月1月的月均温-2.5℃,最热月7月的月均温11.4 ℃。纳帕海为典型的季节性湖泊,雨季湖水上涨,干季湖水通过西北角落水洞,最终汇入金沙江[16]。沿海拔高程,纳帕海在空间上形成湖泊、原生沼泽、沼泽化草甸、草甸等空间分布格局。
纳帕海具有重要的生物多样性与特有性保护价值,分布有全球15种鹤类中唯一生活在高原的国家Ⅰ级保护鸟类黑颈鹤(Grusnigricollis)和黑鹳(Ciconianigra),以及滇西北高原特有鱼类中甸叶须鱼(Ptychobarbuschungtienensis)等。1984年,经云南省政府批准在其主要区域建立纳帕海省级自然保护区;2005年,其核心区域被指定为国际重要湿地,成为国内外关注生物多样性保护的热点区域[16]。近年来,由于当地人口的快速增长以及对湿地资源的依赖与不合理利用,湿地生态系统受到了诸如排水垦殖、过度放牧、基础设施建设等人为活动干扰,以及降水与气温等气候因子变化的影响,湿地生态系统的结构与功能受到较大的威胁[22]。
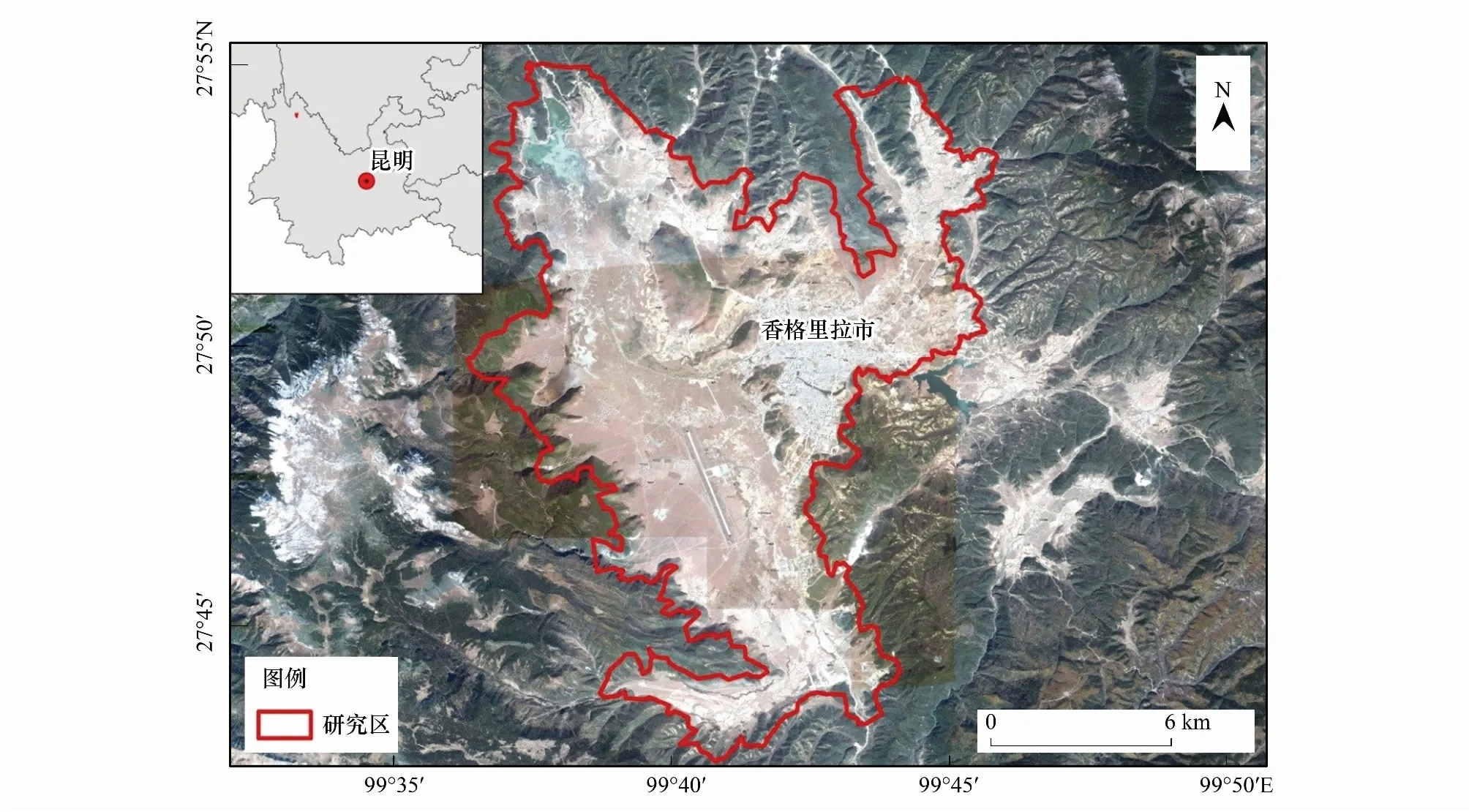
图1 研究区域位置及研究范围Fig.1 Location of study area and study region
1.2 研究技术与方法
1.2.1 景观类型划分及技术方法
基于景观的定义,结合研究区域具体的生态环境特征,以及各自然要素相互作用的关系,以反映景观的内在属性及其差异[23]。依据纳帕海汇水区的海拔高程、水文状况、植被类型及其空间分布,将研究区域划分为湿地类景观和非湿地类景观两大类。其中,湿地景观类型划分为湖泊、原生沼泽、沼泽化草甸、草甸,具体的划分依据见表1。非湿地景观主要包括耕地、建筑用地、林地和灌丛。
1.2.2 数据来源及处理
选取1987年至2015年时段的10期Landsat影像(表2),使用30 m分辨率的 DEM 数据提取纳帕海的汇水区,作为研究范围。影像数据预处理包括影像数据读取-数据辐射校正-几何精校正-图像增强,根据提取的汇水区范围进行图像裁剪。所有的空间数据的投影参考系统为通用横轴墨卡托投影,参考椭球体为WGS84。在确定各年度景观单元的区划之后,在E-Cognition软件支持下,基于遥感影像颜色、纹理、形状,以及数字高程和其他矢量数据进行自动区划,得到形状与图像光谱特征具有同质性的生态景观多边形单元[24]。根据野外遥感建标结果,对各年度生态景观单元进行目视解译分类判读,得到不同年度的生态景观分类结果。2016年11月,运用GPS对研究区域内分类结果进行野外验证及校正,总计验证点达121个,包含研究区域8种景观类型,验证分类精度达85%以上。结合研究区域基础地理数据,绘制研究区域内景观类型图。
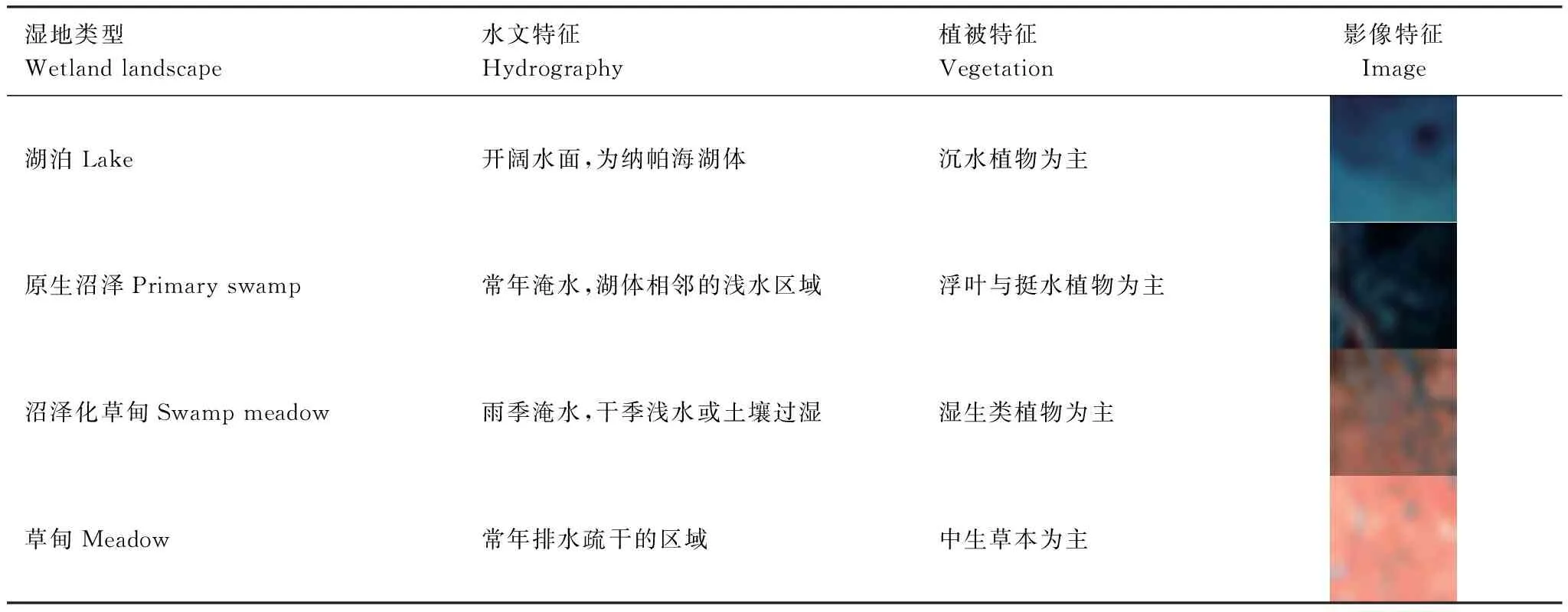
表1 纳帕海湿地景观类型及划分依据
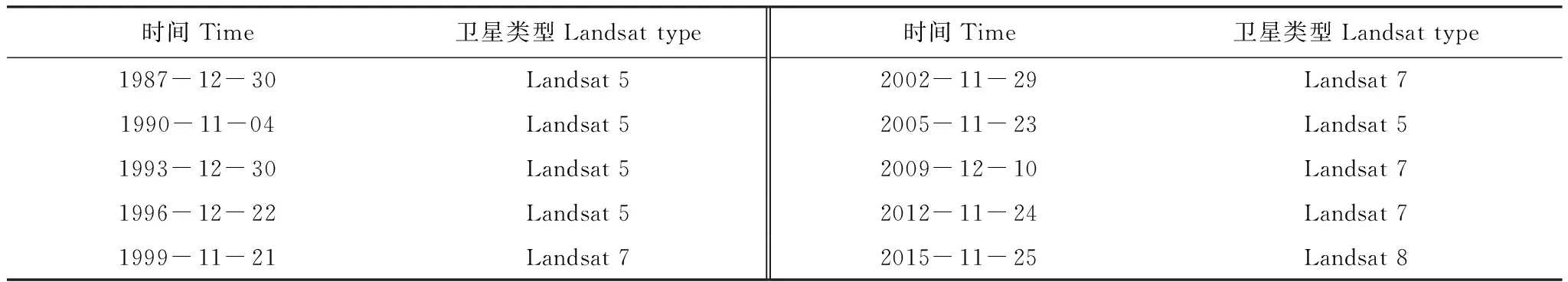
表2 1987年至2015年所用遥感影像的信息
1.2.3 景观格局指数计算
运用ArcGIS 10以及Fragstats 3.4分析研究区域内湿地28年来景观类型组成及其空间格局变化,景观空间格局特征主要选取斑块数目(number of patch, NP)、香农多样性指数(Shannon′s diversity index, SHDI)和集聚度指数(contagion index, CONTAG)以反映研究区域景观多样性特征[25]。
1.2.4 人为与自然因子的收集与处理
区域社会经济发展指标可综合反映人为活动对自然资源干扰的类型和强度[26]。因此,本研究选择社会经济发展的主要经济指标,作为人为活动对研究区域湿地生态系统的干扰指标,主要是第一、二、三产业的经济指标。近30年来,研究区域社会经济总量不断增加,国民生产总值(GDP)由1987年的7956万元增至2015年的585163万元,增长73.5倍。其中,自2006年始,第二、三产业总值呈快速增长,而第一产业总值增长相对缓慢(图2)。
基于该区域自然因子变化特征,以及有关数据的可获得性,本研究选择降水、温度作为自然因子的指标。近30年来,纳帕海地区气温呈现显著增加的趋势,气温上升率为0.43 ℃/10 a(图3);降水量呈现先升高后降低的趋势,降低的趋势在2002年后变为更加明显(图3)。
1.2.5 数据统计分析
运用Sigmaplot 10,分析湿地分布面积、斑块数量、香农多样性指数、聚集度指数与第一、二、三产业产值,以及年均温和降水量的线性关系。运用Canoco 4.2分析软件,通过冗余分析(RDA)方法,分析第一、二、三产业产值,以及年均温和降水量对湿地分布面积、斑块数量、香农多样性指数、聚集度指数的影响差异;并依据偏蒙卡罗特检验来评估每个因子对湿地面积、斑块数量、香农多样性指数、聚集度指数的解释贡献,分析人为活动干扰、自然因子变化对研究区域湿地分布面积和景观多样性变化的影响与作用贡献。
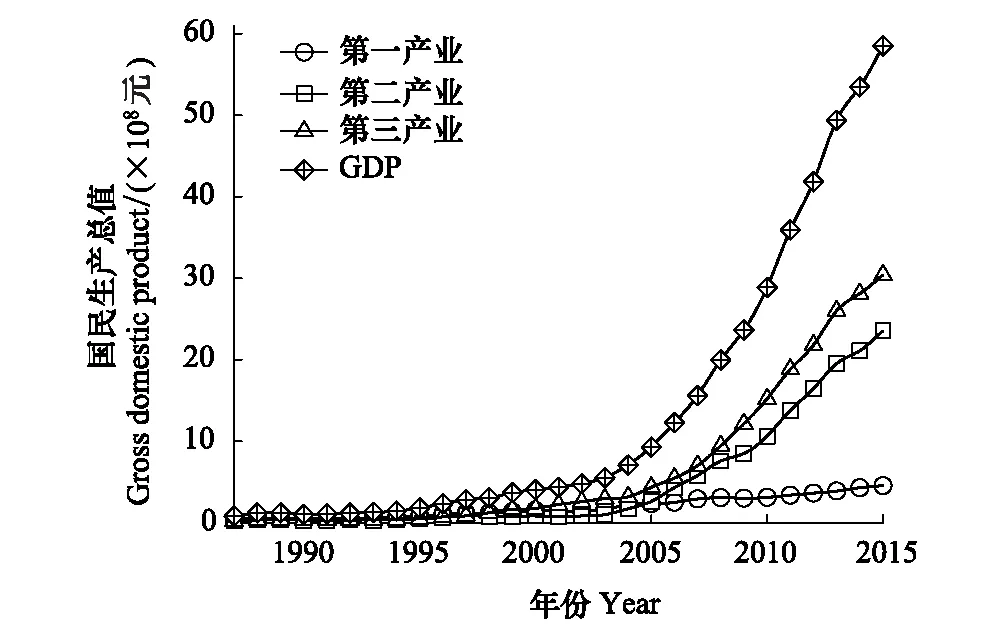
图2 1987—2015年研究区域社会经济发展动态Fig.2 Dynamic of GDPs in study area from 1987 to 2015
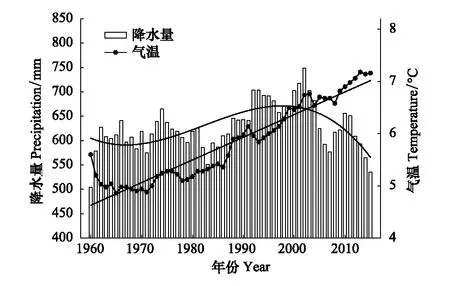
图3 1960—2015年研究区域5年滑动年均气温与年均降水量 Fig.3 Dynamic of annual mean temperature and precipitation in study area from 1960 to 2015
2 结果
2.1 湿地类型及面积变化
在1987—2015年期间,研究区域的湿地面积及其空间分布发生了明显的变化(图4、图5)。研究区域湿地总面积由9194.45 hm2减小为6737.79 hm2,减少率达到26.72%。其中,原生沼泽与沼泽化草甸减少的面积分别为1152.07 hm2和1257.72 hm2(图4);湖泊面积略有增加,由1987年362.30 hm2增加到2015年的518.37 hm2;草甸面积略有减少,由1987年的5649.28 hm2减少到2015年的5446.54 hm2(图4、图5)。
在湿地类型间的转化中,草甸主要来源于湖泊、原生沼泽和沼泽化草甸,共计1449.42 hm2;而草甸转化为湖泊、原生沼泽和沼泽化草甸仅有152.07 hm2。湖泊面积的增加,主要源于原生沼泽与沼泽化草甸,共计211.98 hm2。另外,由湿地类型转换为非湿地类型的面积共计3088.92 hm2,占湿地总面积的33.60%(表3)。
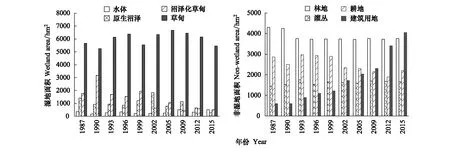
图4 1987—2015研究区域湿地类型与非湿地类型分布面积的变化Fig.4 Changes in wetland types and non-wetland type in study area from 1987 to 2015
2.2 景观多样性及变化
在1987—2005年间,研究区域斑块数量增加,在2005—2015年间其斑块数量下降,最大值为299(2005年)、最小值为200(1990年)(图6)。在1987—1999年间,香农多样性指数呈现增加趋势,而在1999—2015年间却呈现下降的趋势,最大值为1.84(1999年)、最小值为1.75(2015年)(图6)。聚集度指数总体呈先降低后增加的趋势;其中,最小值为52.02(1999年)、最大值为53.85(2012年)(图6)。
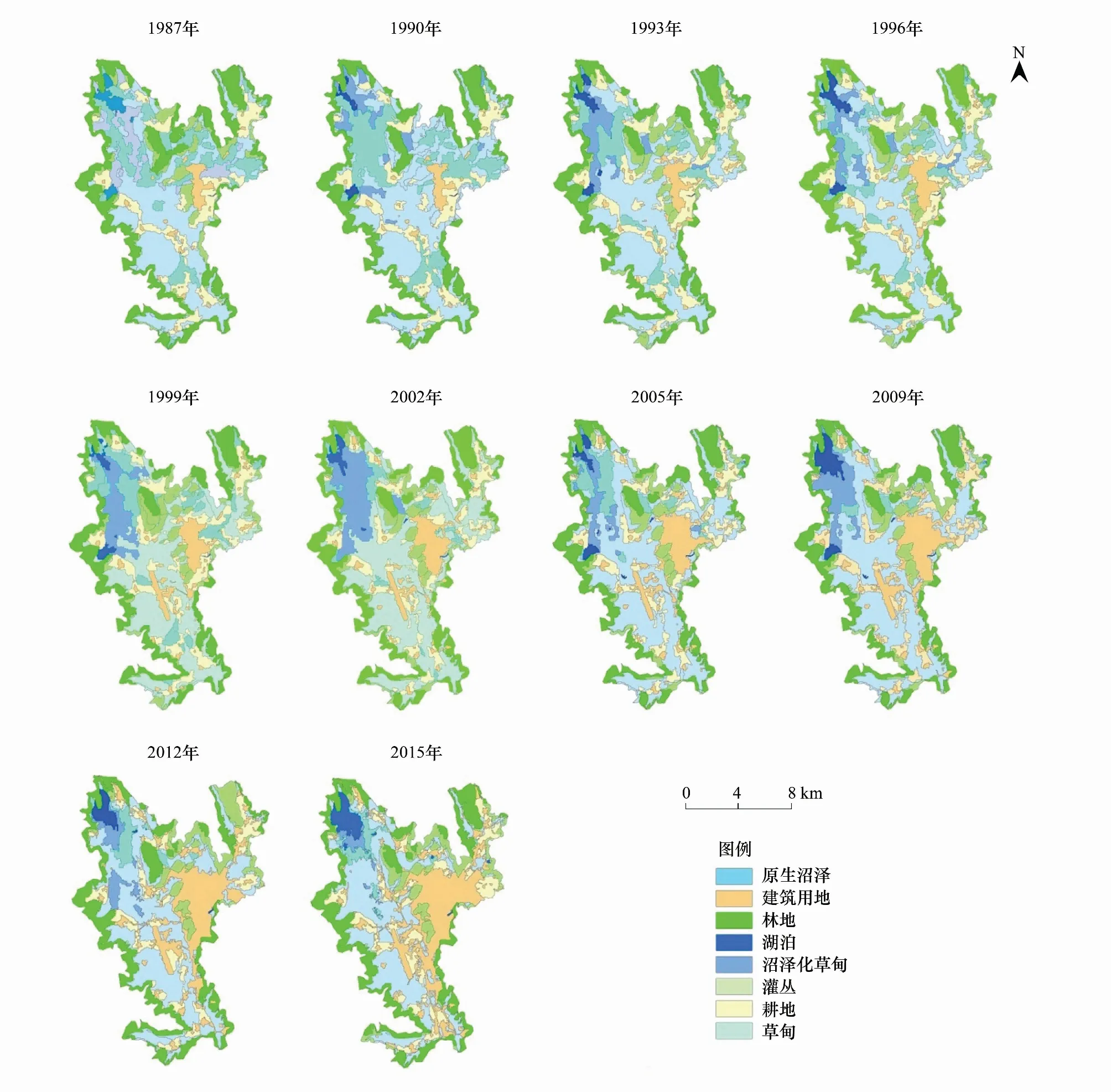
图5 1987—2015年研究区域景观变化Fig.5 Changes in landscapes of study area from 1987 to 2015
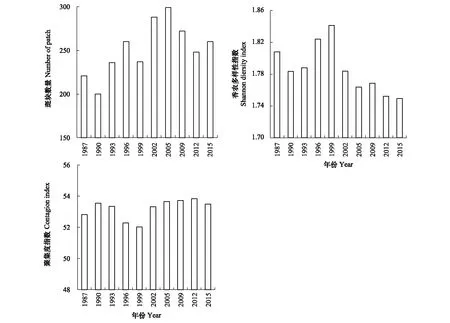
图6 1987—2015年纳帕海景观多样性指数的变化Fig.6 Landscape diversities in the study area from 1987 to 2015
2.3 湿地景观变化与自然、社会经济发展的关系
湿地分布面积与第一、二、三产业呈显著的线性负相关(图7),并随着经济发展指标的增加而呈下降趋势。湿地分布面积与年均温呈显著的线性负相关(图7),与降水呈正相关但相关性不显著(图7)。RDA分析结果表明(图8),湿地与社会经济和自然因子相关性的大小表现为第三产业>第二产业>第一产业>年均温>降水。
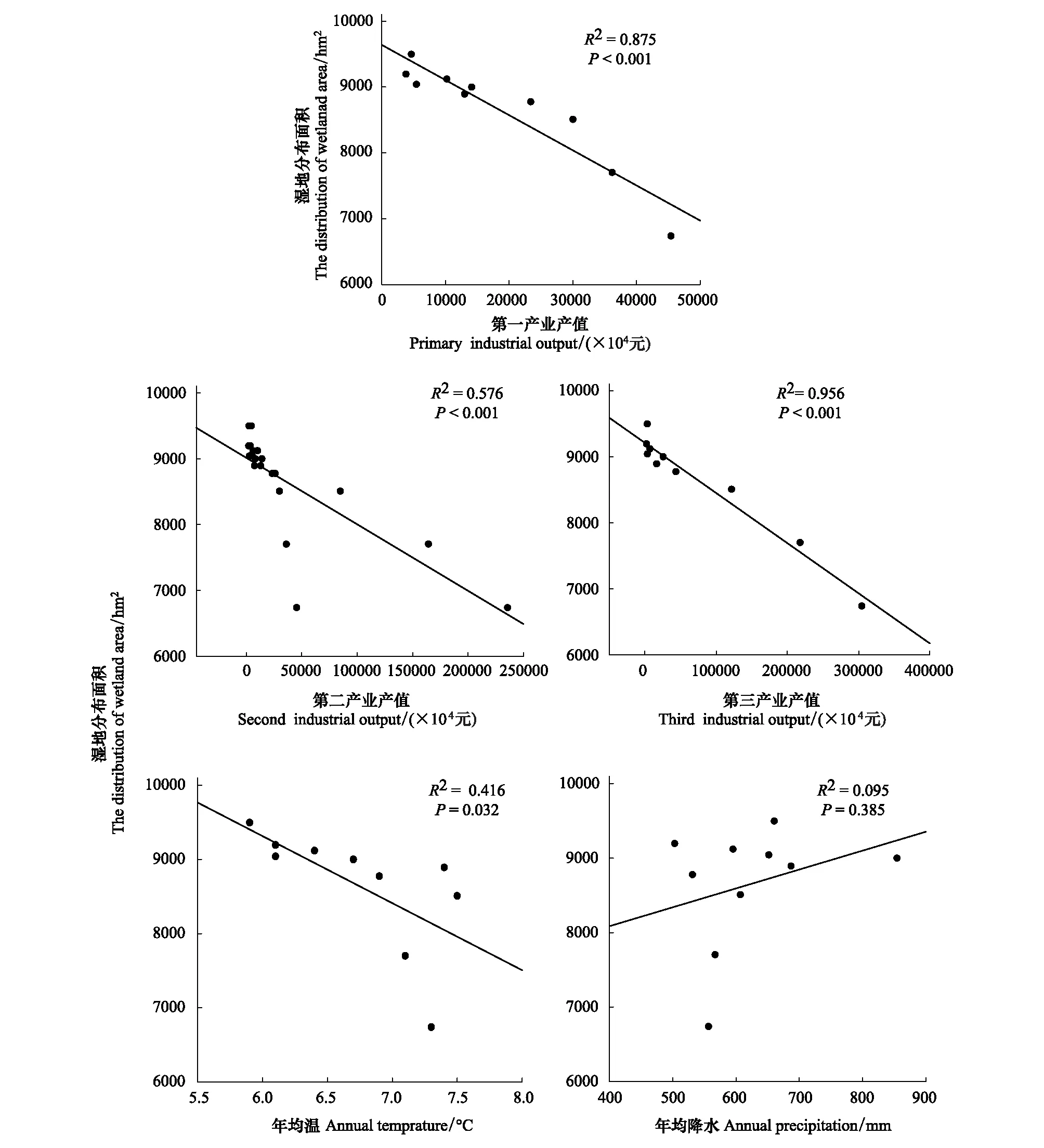
图7 湿地分布面积与主要社会经济和气候指标的相关性Fig.7 Linear relationship linking wetland area and social economic and natural indexes from 1987 to 2015
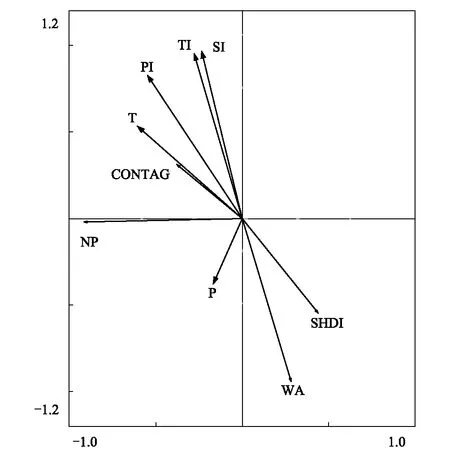
图8 冗余分析 (RDA)下湿地分布及景观多样性与研究区域社会、自然关键因子的相关关系Fig.8 The correlations between wetland distribution, landscape diversity indexes and local social-economic and natural factors revealed by the RDA analysisWA: 湿地分布面积, the distribution of wetland area;NP: 斑块数量,number of patch;SHDI: 香农多样性指数,Shannon Diversity Index;CONTAG: 聚集度指数,the contagion index;PI: 第一产业,the primary industry; SI: 第二产业,the second industry;TI: 第三产业,the third industry;T: 年温度 the annual temperature;P: 年降水,the annual precipitation
斑块数量与第一、二、三产业值呈正相关(图9),但未达到显著性;与年均温(正相关)、降水(负相关)线性相关性不显著(图9)。RDA分析结果表明(图8),斑块数量与社会经济和自然因子相关性的大小表现为年均温>第一产业>降水>第三产业>第二产业。
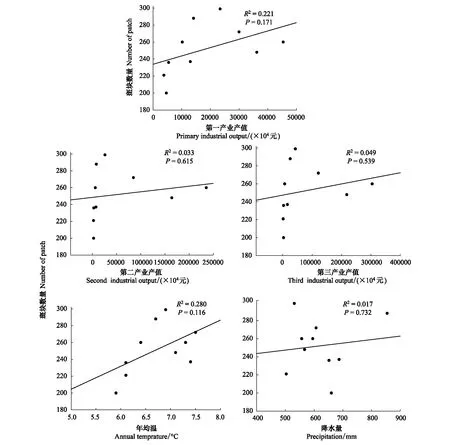
图9 斑块数量与社会经济指与自然因子关键指标的相关性Fig.9 Linear relationship between number of patch and local social-economic and natural factors from 1987 to 2015
香农多样性指数与第一、二、三产业指标呈显著的负相关(图10);与年均温(正相关)、降水(负相关)线性相关性不显著(图10)。RDA分析结果表明(图8),香农多样性指数与社会经济和自然因子相关性的大小表现为第一产业>年均温>第三产业>第二产业>降水。
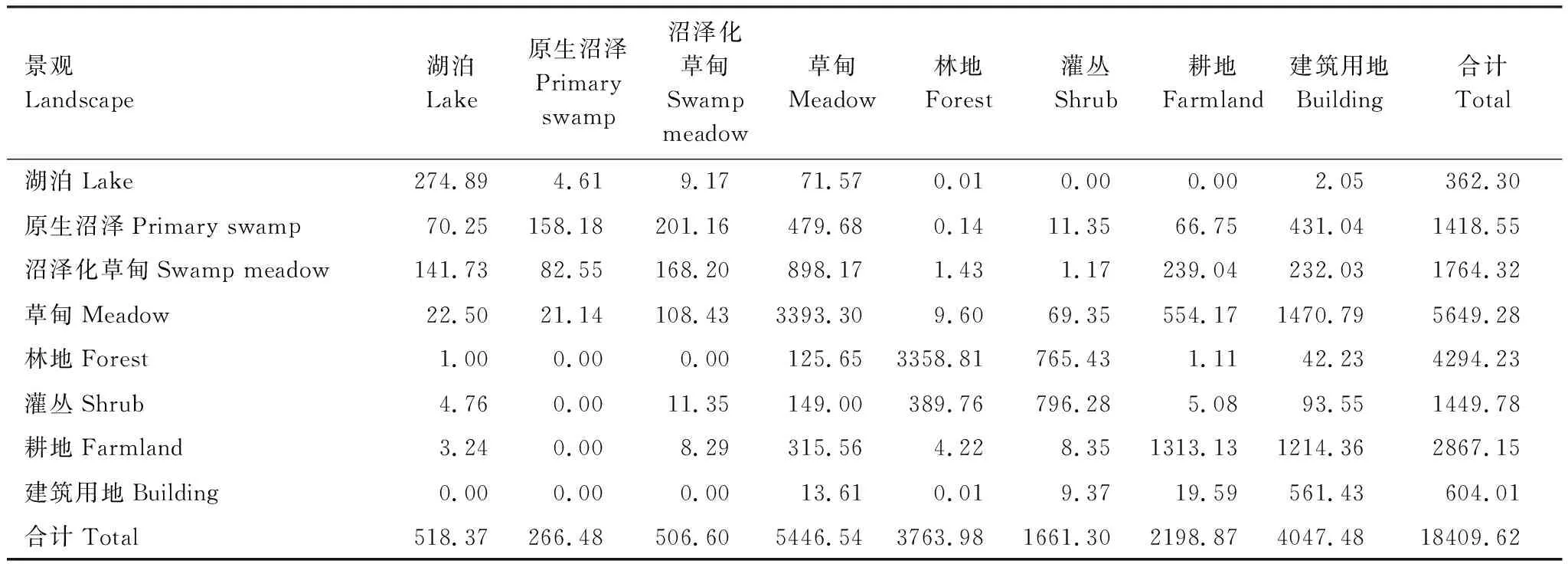
表3 1987—2015年湿地景观类型面积转移矩阵/hm2
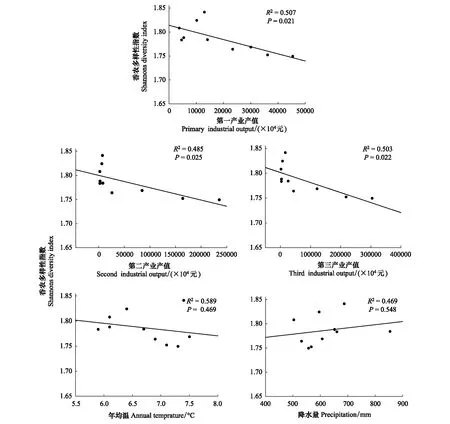
图10 香农多样性指数与社会经济、年均温、年均降水的相关性Fig.10 Linear relationship between Shannon′s Diversity Index and local social-economic and natural factors from 1987 to 2015
聚集度指数与第一、二、三产业值呈正相关,但未达到显著性(图11);与年均温(正相关)、降水(负相关)线性相关性不显著(图11)。RDA分析结果表明(图8),香农多样性指数与社会经济和自然因子相关性的大小表现为年均温>第一产业>第三产业>第二产业>降水。
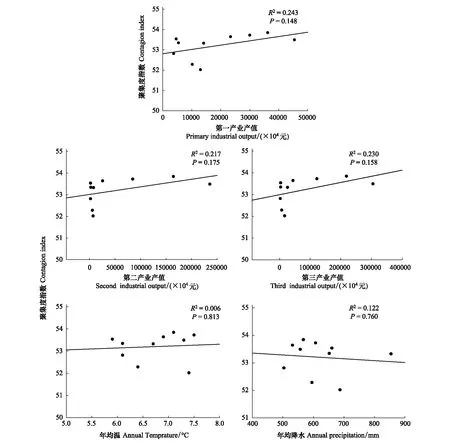
图11 聚集度指数与社会经济、年均温、年均降水的相关性Fig.11 Linear relationship between contagion index and local social-economic and natural factors from 1987 to 2015
RDA分析表明,第一、二、三产业对湿地面积和景观多样性变化的影响权重大于年均温和降水,其中,社会经济对湿地面积、景观多样性变化整体的解释度为63.50%,气候因子对其的解释度36.50%(图8)。
3 讨论
3.1 人为活动干扰与湿地景观
本研究表明,近30年来研究区域湿地分布总面积呈现减少的趋势,斑块数量总体呈现先增加后减少的趋势,香农多样性指数也呈先升高后降低的趋势,聚集度指数呈先降低后升高的趋势。
湿地面积的减少和景观多样性的变化趋势与本区域社会经济发展的关键指标密切相关,其中,湿地面积和香农多样性指数与第一、二、三产业值、年均温均呈负相关,与降水呈正相关;斑块数量和聚集度指数与第一、二、三产业、年均温均呈正相关,与降水呈负相关。综合来看,社会经济因子对湿地面积和景观多样性的解释度为63.50%,自然因子的解释度为36.50%。
纳帕海草甸、沼泽化草甸是香格里拉天然的牧场,放牧成为该地区第一产业值中的主体。近30年来香格里拉牧业产值从1987年到2015年,增加了22639万元,增加率达1524.51%。随着牧业经济快速发展,过度放牧成为影响该区域湿地的重要人为干扰活动。据研究,纳帕海地区理论载畜量为1567黄牛单位,而当前实际载畜量(主要以牦牛、马和猪统计)为7486黄牛单位,超载率377.73%[12]。过度放牧通过牲畜的采食、践踏影响该地区植物的生长、发育,以及种间关系;另外,过度放牧也致使土壤容重的增加,含水量的降低,土壤养分的降低以及湿地生境不断丧失,进而影响原有湿地植物的栖息生境,导致原有植物群落结构改变,直接影响湿地景观的类型与空间分布。自2009年该区域藏香猪放牧超载率达300%[27],藏香猪在觅食过程中不断拱翻土壤,形成大片条状、带状或片状的裸露土地,不断改变湿地植被组成及其空间分布,也影响了湿地景观。
排水垦殖为主的种植业发展,是第一产业中改变湿地景观的另一重要因素。近30年来,研究区域种植业产值从1987年的1311万元增长到2015年的30778万元。由于湿地区居民对湿地的排水垦殖活动,使得湿地分布面积不断萎缩,湿地的类型及空间分布格局随之发生变化[28]。另外,不断增加的农业灌溉用水也使得流入湿地的水量减少,导致湿地分布面积萎缩。过度放牧、排水垦殖等活动,直接导致小斑块数量减少、大斑块面积增大,即景观的空间异质性降低、同质性增加,进而导致整个区域景观的类型多样性降低、集聚度指数增大。
1987年到2015年,研究区域第二产业值得到快速发展,从2241万元增加到235510万元。在第二产业快速增长的过程中,其主要依托是不断扩张的香格里拉市区及基础设施建设。研究表明,整个城市建筑面积从1987年的604.01 hm2增长到2015年的4047.48 hm2。快速的城市发展,直接导致了湿地分布面积的减少、类型的改变,以及空间分布格局的改变。在研究的前期(1987—1999年),城市的扩展与基础设施建设相对较慢,对建筑用地面积的需求相对较小,但到了后期(1999年—2015年),城市与基础施设建设不断增强,导致空间异质性降低、同质性增加,进而导致景观类型降低、聚集度指数增加。
另外,随着香格里拉旅游业的快速发展,研究区域湿地成为了香格里拉重要的旅游景点。自1987年到2015年,第三产业值由1954万元增长到304257万元。旅游生活污水的排放加剧了纳帕海水体富营养化的过程,从而改变植物的分布格局;为了发展旅游业而修建的环湖公路影响湿地的水分补给过程,改变湿地原有的生境;无序旅游中马匹和人的过度践踏导致土壤表层物理性状改变,增加土壤紧实度,破坏了沼泽土和泥炭土的草根层,加速了有机质分解,促进了矿化作用,导致沼泽土壤养分含量衰减下降,影响了植物生长的土壤环境条件;践踏作用也使植被稀少的面积增加,植物高度降低,践踏也使原有的物种被耐磨物种取代[16],导致物种结构和多样性的改变,进而改变湿地景观格局。
3.2 自然因子的变化与湿地景观
本研究中,温度与降水的变化对整个研究区域湿地类型、分布面积及景观多样性存在影响,但解释度相对较小。通常情况下,温度和降水变化可通过对湿地植物的生长、种间关系,土壤养分条件,湿地水文等直接与间接的作用,进而影响湿地分布面积、类型及空间分布格局。相关研究表明,在降水不足的情况下,湿地分布面积明显萎缩[29],植物的生长受到抑制[30],也导致土壤由淹水的厌氧环境进入有氧环境,使土壤表层处于较干燥的氧化环境而利于有机质矿化[31],影响植物的养分供给和生长,导致湿地地表景观类型的最终变化。也有研究表明,气温升高对该区域湖滨带优势植物水葱(Scirpustabernaemontani) 和茭草(Zizaniacaduciflora)的生长有促进作用,而对刘氏荸荠(Heleocharisliouana)和小黑三棱(Sparganiumsimplex)的生长产生胁迫[32]。这种物种对气温上升的差异响应,将导致种间关系的变化,从而使得原有的湿地植被空间分布格局变化,最终影响湿地的景观类型及其多样性。
`3.3人为活动的干扰与自然因子的变化对湿地景观作用的差异
本研究表明,人为活动的干扰和自然因子的变化对研究区域湿地面积和景观变化的解释度不同。其中人为活动是导致纳帕海湿地分布面积减少、空间格局变化的主要驱动因子,这与其他地区研究结果相似[33- 34]。
解释度的差异主要与人为干扰与自然变化对湿地生态系统的作用特征密切相关。通常情况下,人为活动对湿地的干扰主要作用于较小的空间尺度以及时间尺度,具有途径多样、强度高、速度快、持续时间相对较短等特点;而自然因子的影响主要在较大的时间尺度以及空间尺度上,具有影响范围大、持续时间长,时滞性等特征[35]。本研究中,由于研究区域范围有限时间跨度相对较短,人为活动干扰通过快速改变土地利用方式影响湿地景观特征,本研究区域仅城市建设面积就增加了近5.70倍。而自然因子通常需要通过改变湿地水文过程,影响土壤养分,改变湿地植物的生长、发育以及湿地植物群落组成、种间关系等,进而对湿地类型及景观造成影响,其过程具有明显的滞后性[30]。因此,在较小时间和空间尺度上,相对于人为活动的干扰,自然因子的变化对湿地生态系统影的贡献相对较小。
4 结论
人为活动的干扰、自然因子的变化,共同影响着滇西北高原湿地的类型、空间分布及景观多样性,但两者对湿地生态系统的作用贡献存在差异。人为活动干扰对湿地面积、景观多样性指数的解释度为63.50%,自然因子变化的解释度36.50%。研究进一步表明,近30年来,人为活动的干扰是影响滇西北高原地区湿地生态系统的关键驱动力,减缓人为活动对湿地生态系统的过度影响,是实现当地湿地保护的关键。
猜你喜欢
小学生学习指导(高年级)(2022年5期)2022-06-02 05:16:08
数学小灵通·3-4年级(2021年5期)2022-01-01 08:00:14
黑龙江气象(2021年2期)2021-11-05 07:07:00
中学生数理化·中考版(2021年8期)2021-07-31 07:41:44
摄影之友(影像视觉)(2019年2期)2019-03-05 08:27:20
中学生数理化·七年级数学人教版(2018年4期)2018-06-28 03:26:30
家教世界(2018年16期)2018-06-20 02:22:00
中国篆刻(2017年5期)2017-07-18 11:09:30
成都信息工程大学学报(2016年6期)2016-06-01 12:10:06
Acta Mathematica Scientia(English Series)(2015年6期)2015-02-10 08:37:17
