
不同生态型芦苇种群对盐胁迫的生长和光合特性
2019-02-20 10:43:04许振伟宋慧佳李明燕张廷靖FRANZISKAEllerHANSBrix侯文轩郭卫华
生态学报 2019年2期
许振伟,宋慧佳,李明燕,张廷靖,郭 霄,2,FRANZISKA Eller,HANS Brix, 杜 宁,侯文轩,郭卫华,*
1 山东大学生命科学学院, 青岛 266237 2 青岛农业大学园林与林学院, 青岛 266109 3 丹麦奥尔胡斯大学,生命科学学院植物科学系,奥尔胡斯 8000
随着全球气候变暖问题的加剧,干旱和半干旱地区的蒸发量和矿物质积累增加[1],湿地土壤盐渍化和次生盐渍化面积逐渐扩大,成为各国关注的全球性问题[2]。我国土壤盐渍化分布面积大,范围广,主要包括内陆的盐渍化和沿海土壤盐渍化两种类型。采用生态措施改善盐渍土是目前土壤盐渍化改良的重要方向,能够极大地提高盐渍土壤的利用效率[2]。
芦苇(Phragmitesaustrilis(Cav.)Trin.ex Steud.)是一种具有广泛生境适应能力的水生挺水物种,其繁殖能力极强,生态幅极广,可在滨海滩涂、河流沿岸、河口等浅水湿地形成典型的单优群落,而且可以在盐碱、荒漠地区也可广泛分布,有着极好的耐盐特性[3]。研究发现,在盐处理条件下,芦苇可以通过增强吸水,增强细胞膜的稳定和膜功能,促进芦苇生理生化代谢,进而促进幼苗的生长和生物量积累,来提高其耐盐性[4]。同时,芦苇通过调控渗透压的有关物质和降低超氧化物阴离子产生速率降低盐分对于植物生长的毒害作用[5]。研究表明,芦苇在适应盐胁迫有着较好的潜力[6],可以在中等盐渍化的土壤中正常生长[4],成为改良盐渍化土壤一种可行性的材料。
不同生境下的芦苇种群在长期的进化过程中,形成了生态学上互有差异的类群,具有各自稳定的形态、生理生化和生态学等特征,构成丰富的生态型[3]。不同生态型的芦苇种群因其所处的环境不同,在进化的过程中其形态、生理特征等都发生了不同程度的变化,来应对各种形式的环境压力[7]。Li等人比较研究了陆生型芦苇与水生型芦苇功能性状的差异,发现两者主要是通过叶片和根的功能性状改变来适应环境的变化,两者有着不同的资源分配策略,水生型芦苇主要是通过调整叶面积和根的长度进行快速的生长,但是对环境胁迫敏感,而陆生型芦苇通过调整叶片和根中的干物质含量对变化的环境进行适应,使得其自身生长缓慢,但同时也造就了其对环境胁迫具有高耐受性的优势[8]。
芦苇生长所需要的能量主要来自于光合作用固定的光能,但是盐胁迫却会对芦苇的光合作用产生消极影响[9]。盐胁迫造成芦苇光合作用下降主要包括两部分原因,一是气孔因素,低程度盐胁迫会造成芦苇气孔关闭,使得芦苇光合作用底物供应不足[10];二是非气孔因素,严重盐胁迫造成芦苇光合结构和功能损伤,进而造成芦苇光合作用的下调[7,9]。尽管国内外学者对芦苇光合作用对盐胁迫响应机制有部分的研究,但是不同生态型的芦苇的光合作用对盐胁迫的响应研究却很少[1],因此本实验将不同生态型的芦苇的光合作用作为研究重点,通过模拟滨海盐渍化土壤,比较内陆型与河口型芦苇在含盐(2.00%)与无盐(0.00%)的处理下的生长及光合特性,对比不同生态型芦苇种群的耐盐机理,并推测两种生态型芦苇可能存在的耐盐机理差异的原因,为盐渍化土壤的改良提供科学依据。由于河口型芦苇长期生活在盐渍化土壤环境中,我们预测,较之内陆型芦苇,长期的生态适应将使得河口型芦苇拥有更强的耐盐性。
1 材料与方法
1.1 材料与装置
实验于2017年4月—8月在山东省莱芜市房干野外实验基地种植园(36°26′N,117°27′E)中进行,为典型的温带季风型气候,四季分明,雨热同期,夏季高温多雨,冬季干旱少雨,光、热、水等条件较优越,年平均气温为12.4℃,≥10℃年积温为4200—4600℃,年降水量为600—830 mm[11]。内陆型(CN)共11种类型采于内陆,每种标记为1个类型,河口型(YRD)共11种类型采于黄河三角洲,每种标记为1个类型(表1)。对于同一生态型的芦苇,在每个采集点选择生长状况良好且较为一致的芦苇作为采集样本,采集芦苇根系至实验室进行栽种培养。在第二年,每种芦苇剪取若干且粗细长度一致的根状茎作为繁殖体进行移栽,每个容器内种植一段。
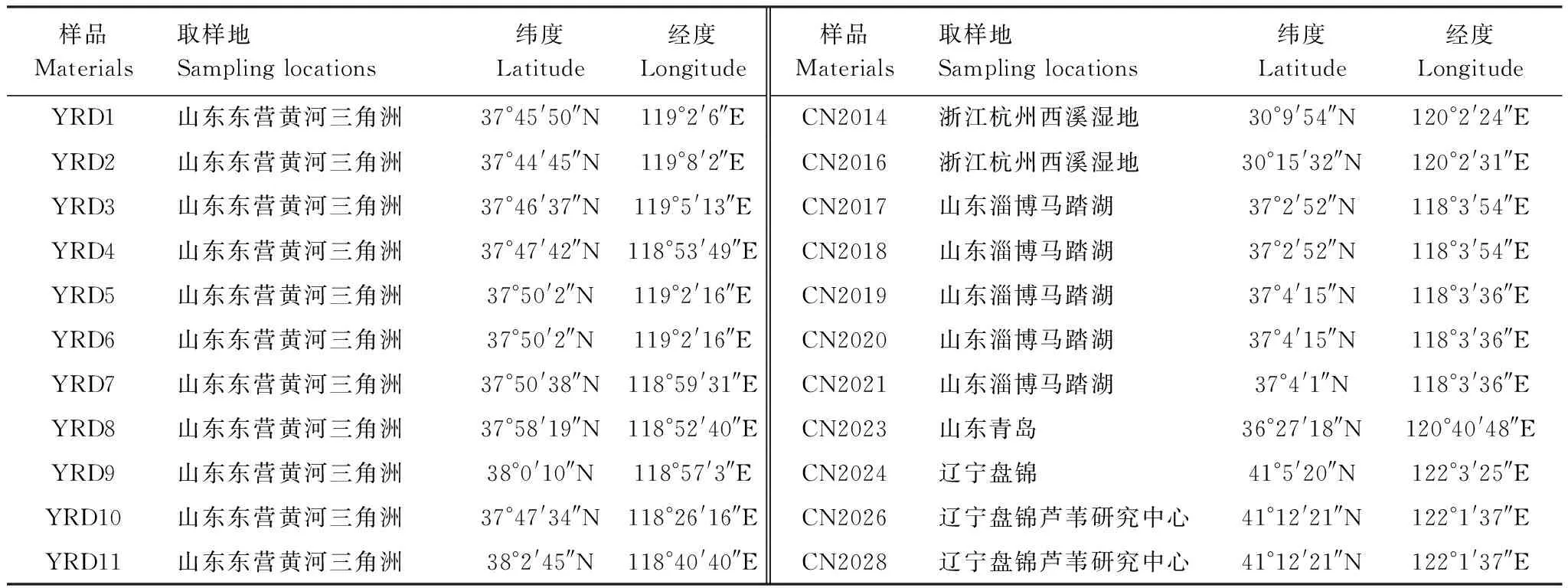
表1 芦苇群落取样地点
YRD:黄河三角洲,Yellow River delta;CN:中国内陆
1.2 实验设计
实验时间为2017年4月—8月,将芦苇的盐度设置为淡水处理(0.00%),含盐处理(质量浓度2.00%)共两个盐度梯度,0.00%梯度使用淡水进行灌溉,2.00%梯度,在种植3—4周后进行加盐处理,首次加盐至盐质量浓度1.00%,后两次加盐每次提高0.50%盐浓度,每次加盐间隔1周。芦苇种植于20 L的塑料桶中,土壤基质为河沙,种植时添加缓释肥料(约50 g/盆)和0.5 g/盆络合铁(Fe-EDDHA)将其混河沙中,自来水充分浸透沙子并且沙子表面无积水。每个处理随机分布在试验小区内,并且在每个地块内围成一个面积为1 m2(1 m×1 m)的样地。各个处理都保持灌溉水层为30 cm。
1.3 测量方法
在芦苇生长初期(5月初)、成熟期(8月末) 在每个地块内选出长势均一的5株芦苇测量其株高(height),测量测量5个最高的芽时自盆土顶部到顶端(包括顶端叶)的高度,选取实验开始处理时芦苇的株高数据H1和收获时株高数据H2,相差时间为ΔT,用于计算相对生长速率(RGR);在芦苇的生长末期测量每盆株数(shoot number)。每个处理选择两株测量叶片的鲜重(FW),干重(DW),饱和重(TW),计算叶片相对含水量(RWC)。选取晴朗天气,使用CCM- 200(Opti-Sciences, USA)叶绿素含量测定仪测定芦苇功能叶片叶绿素总含量(chla+b),叶绿素比(chla/b);在收获日9:00—11:00,使用LI- 6800(Li-Cor, USA)便携式光合仪测定芦苇功能叶片的净光合速率(A),气孔导度(Gs),蒸腾速率(E),计算水分利用效率(WUE)。
其中:RWC=(FW-DW)/(TW-DW);
RGR=InH2-InH1/ΔT;
WUE=A/E。
1.4 数据分析
测量的数据使用Excel 2013进行数据的整理,使用SPSS for windows 13.0(SSPS Inc, USA)数据分析软件进行双因素方差分析(Two-way ANOVE)检验不同盐度、生态型和两者交互作用的影响和单因素方差分析(One-way ANOVA)检验不同处理间的差异,并采用Turkey HSD法进行多重性比较,
用字母标记法在各图中标出统计分析结果,绘图制作工具采用Origin 8.0(Origin Lab Co, Northampton, MA, USA)。
2 实验结果
2.1 盐胁迫对两种生态型的芦苇生长指标的影响
在实验过程中观察发现,尽管河口型和内陆型芦苇虽然采集于不同地点,但是同一生态型的芦苇生长状况较为一致。由图1可知,在加盐处理下,河口型芦苇的株高为120.5 cm,与淡水处理相比,下降33.0%;内陆型芦苇的株高为124.7 cm,下降32.1%,盐胁迫显著性的降低了两种生态型的芦苇株高(P<0.05)。在淡水处理和加盐处理条件下,两种生态型的芦苇的株高均没有显著性差异(P>0.05)。在加盐处理下,与淡水处理相比,河口型芦苇和内陆型芦苇的相对生长速率分别下降33.7%和37.9%,降低显著(P<0.05)。在淡水和加盐处理条件下,河口型芦苇的生长速率显著性地高于内陆型芦苇(P<0.05)。盐胁迫并没有对芦苇的株数和叶片相对含水量产生显著性影响(P>0.05),两种生态型芦苇的株数在相对含水量在淡水和加盐处理下并没有显著性差异(P>0.05)。
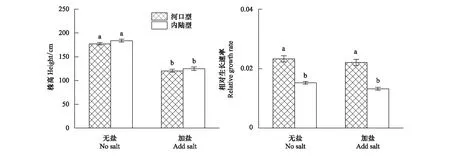
图1 不同品系芦苇在不同盐度下生长指标的变化情况(数值为平均值±标准误差)Fig.1 The growth parameters of different ecotypes reeds in different salinities (mean ± SE)
在双因素方差分析中, “*”代表不同组别间差异显著,P<0.05; “**”代表不同组别间差异极显著P<0.01
2.2 盐胁迫对两种生态型的芦苇光合指标及叶绿素指标的影响
植物的光合作用和蒸腾作用是对胁迫的直观体现。在加盐处理下,与淡水处理相比,河口型芦苇和内陆型芦苇的净光合速率分别降低19.7%和16.7%,降低并不显著(P>0.05),在淡水处理下,河口型芦苇的净光合速率显著性高于内陆型芦苇(P<0.05),但在加盐处理下,两种生态型芦苇的净光合速率没有显著性差异(P>0.05)。盐胁迫造成两种生态型芦苇的蒸腾速率显著性降低(P<0.05),无论在淡水还是加盐处理条件下两种生态型芦苇的蒸腾速率没有显著性差异(P>0.05)。在加盐处理条件下,与淡水处理相比,两种生态型芦苇的水分利用效率显著性提高(P<0.05)。在加盐处理下,河口型芦苇的气孔导度为434.2042 mol m-2s-1,但与淡水处理没有显著性差异(P>0.05),但在加盐处理下,内陆型芦苇的气孔导度显著性下降(P<0.05),两种生态型芦苇的气孔导度存在显著性差异(P>0.05)。
在淡水和加盐处理下,两种生态型芦苇的总叶绿素浓度均维持在400 mg/g以上,叶绿素比在1.25以上,与淡水处理相比,盐胁迫使得河口型芦苇的总叶绿素浓度和叶绿素比略有上升,内陆型芦苇的总叶绿素浓度和叶绿素比略有下降,但均不显著(P>0.05)。
3 讨论及分析
3.1 盐胁迫对两种生态型芦苇的影响
从图1得出,与淡水处理相比,在加盐(2.00%)处理条件下,河口型芦苇和内陆型芦苇的株高呈现显著下降趋势(P<0.05),河口型芦苇株高下降33.0%和,内陆型芦苇株高下降32.1%,盐胁迫对两种生态型的芦苇的生长产生显著抑制。我们推测原因可能包括两方面:一方面,芦苇的生长与其净光合速率密切相关[12],盐胁迫会造成芦苇的净光合速率显著性下降,使得芦苇固定光能和碳供应能力明显下降,芦苇的生长所需要的能量和物质供应受到抑制[9],使得两种生态型的芦苇株高下降;另一方面,可能是在盐胁迫下,芦苇将活跃的分生组织生长分裂的能量转移到保持植物渗透平衡中去[13],使得芦苇的茎和新叶的生长受到盐胁迫的抑制[1],造成两种生态型芦苇株高的降低。
净光合速率是反映植物遭受盐胁迫程度和鉴定植物耐盐性的重要指标,直接反映了植物光同化能力[14]。盐胁迫使得两种生态型的净光合速率略有下降,河口型芦苇和内陆型芦苇的净光合速率分别下降19.7%和16.7%,但两者并不显著(P>0.05)(表2,图2),表明两种生态型芦苇的净光合速率有着较高的耐盐阈值,因此我们推测在盐胁迫下净光合速率并不是两种生态型芦苇生长受到抑制的原因。但是,盐胁迫造成两种生态型的芦苇蒸腾速率显著性地下降(P<0.05),河口型芦苇的蒸腾速率下降38.4%,内陆型芦苇的蒸腾速率下降38.6%(表2,图2),在盐胁迫下,芦苇经常出现生理性“干旱”[7],芦苇的根尖和幼嫩叶片细胞膨压降低,进而造成气孔关闭,使得蒸腾作用下调,造成芦苇碳积累不足,抑制芦苇生长[7]。
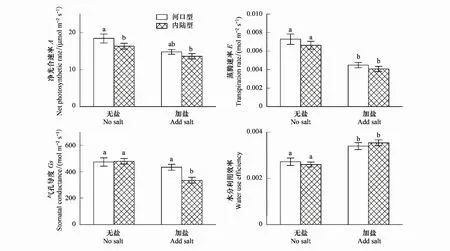
图2 不同品系的芦苇在不同盐度下光合作用指标变化情况(数值为平均值±标准误差)Fig.2 The parameters of photosynthesis of different ecotypes reeds in different salinities (mean ± SE)
水分利用效率是芦苇光合作用和蒸腾作用特征的综合体现,水分利用效率高的植物,叶片保持水分能力强,对水分输出节约,限制叶片表面的水分蒸发,减轻了盐胁迫对植物光合作用的限制,提高了植物对盐胁迫的抗性[15]。在本实验过程中,盐胁迫会造成两种生态型芦苇水分利用效率显著性提高(P<0.05),河口型芦苇的水分利用效率提高24.8%,内陆型芦苇的水分利用效率提高36.1%(表2,图2)。在实验中,盐胁迫并未对两种生态型芦苇的净光合速率产生明显影响,但是却明显降低了两种生态型芦苇的蒸腾速率(图2),明显提高两者水分利用效率,说明适当的盐浓度会显著促进两种生态型芦苇的水分利用效率,该结果与张晓晓等人研究Y65,Y1品系白榆在50 mmol/L盐浓度时水分利用效率结果一致[15]。研究发现,芦苇体内的渗透性调节物质,如脯氨酸,蔗糖含量都会明显提高[16],可溶性蛋白质含量增加[17],提高了两种生态型芦苇的水分利用效率,保证了其正常生长和生理过程所需要的水分。
图3 不同品系芦苇在不同盐度下叶绿素指标的变化情况(数值为平均值±标准误差)Fig.3 The chlorophyll parameters of different ecotypes reeds in the different salinities (mean ± SE)
3.2 河口型芦苇与内陆型芦苇耐盐性差异
在淡水和加盐(2.00%)处理下,两种生态型芦苇的株高没有显著性差异,但是河口型芦苇的相对生长速率显著高于内陆型芦苇(表2,图1)。庄遥提出,同一形态指标的在不同芦苇种群间变化较大,说明芦苇是形态高度变异的物种,这一发现也与本实验不同生态型芦苇的相对生长速率变化相符[4]。
在淡水条件下,河口型芦苇的净光合速率显著性高于内陆型芦苇(P<0.05)(表2,图2),高的净光合速率积累充足的光合产物,保证了芦苇碳供应,使得芦苇有着足够能量进行生长,因此,高的净光合速率可能是在淡水条件下河口型芦苇的相对生长速率明显高于内陆型芦苇(图1,图2)的原因之一。
盐胁迫对芦苇的作用是多方面的,其作用程度对不同生态型的芦苇也有所差异,研究表明,芦苇的在盐胁迫条件下,在较低和中等的盐浓度下,盐胁迫作用于光合作用,造成植物产生应急性反应,像气孔的关闭;在较高的盐浓度下,光系统PSⅡ的供体和受体是盐离子的主要靶点[7],Na+和Cl-对叶绿体造成了一定程度的损伤[18],盐胁迫严重损害了光合作用的结构和功能[1]。
在盐胁迫下,高水平的气体交换是河口型芦苇应对盐胁迫的一种重要策略[12,19]。气孔关闭是芦苇在盐胁迫下对水势降低第一反应,水势降低驱动芦苇对盐胁迫的生理响应,但同时限制了物质的吸收,其中也包括了光合作用的底物[19]。在本实验中,与淡水处理相比,在加盐(2.00%)处理时,内陆型芦苇的气孔导度显著性(P<0.05),降低30.0%,河口型芦苇的气孔导度降低8.2%,但并不显著性(P>0.05),同时,在加盐(2.00%)处理条件下,河口型芦苇的气孔导度明显高于内陆型芦苇(P<0.05)(表2,图2)。植物可以通过调节气孔开放程度,平衡水分散失和CO2交换,达到最适的生理状态[20]。研究表明,在含盐条件下,盐胁迫会使芦苇的水势降低,造成水供应“赤字”,使得芦苇吸收水分的代价增大,为了避免因蒸腾作用丧失过多水分,芦苇会关闭气孔,造成内陆型芦苇的气孔导度降低[18]。Rajput研究盐胁迫对胡杨幼苗影响时也发现,盐胁迫会造成胡杨严重水分胁迫,随着盐浓度的逐渐升高,胡杨幼苗气孔导度逐渐降低,胡杨幼苗通过减小叶片的气孔开口面积和增加气孔密度来避免盐胁迫下过多的蒸腾耗水[21]。在本实验中,在加盐(2.00%)处理条件下,河口型芦苇有着内陆型芦苇更高的气孔导度,保证了其高的气体交换速率,使河口型芦苇光合作用的底物CO2充足,保证高的净光合速率,降低碳匮乏的可能性。
叶绿素是植物叶绿体类囊体膜上重要的光能吸收和转化分子,是植物光合结构的重要组成部分,与植物忍受逆境能力和光活力强弱有密切关系[22]。从表2和图2可以看出,在淡水和加盐处理条件下,同一生态型芦苇的总叶绿素含量和叶绿素比都没有显著性差异(P>0.05)(表2,图3)。同时,Tho等人在研究不同来源的芦苇差异性时也发现,欧洲芦苇种群与北美芦苇种群在同一处理下时叶绿素a,b及叶绿素总量都没有显著性的变化[23]。在种植过程中,在加盐处理一段时间后,尽管两种生态型的芦苇下部叶片出现枯黄现象[24],但是两种生态型芦苇的净光合速率、总叶绿素和叶绿素比并没有显著性下降,说明两种生态型芦苇的光合结构和功能并没有发生明显损伤[18],因此,我们推测叶绿素在芦苇正常生命活动和生长的过程中扮演着重要的角色,是芦苇捕获和转化光能的重要物质,对于维持芦苇高水平的光合作用十分重要。
芦苇在长期的生态适应过程中,不断进化,逐渐形成了适应当地生境的生态型。本研究中,内陆型芦苇与河口型芦苇的初始植株大小、所处环境、处理因素均完全一致,因此不同盐分条件的表型性状的差异是由于植物的表型可塑性导致的,而不同生态型芦苇表现出的表型性状的差异来自于其遗传背景的不同[25]。植物的遗传背景是由于其原产地的环境特征所决定的[26],所以内陆型芦苇与河口型芦苇的耐盐性差异的来源是其原产地生境的不同。
高的相对生长速率能够帮助芦苇迅速占据有利的位置,在应用到实际盐渍化土壤改良中,河口型芦苇有着较高的耐盐能力,通过快速的生长和扩展占领栖息地,成为该地的优势种,起到良好的覆盖作用,进而降低地表的水分蒸发,缓解当地的土壤盐渍化现象。
猜你喜欢
现代装饰(2022年5期)2022-10-13 08:47:08
现代装饰(2020年8期)2020-08-24 08:23:52
重庆与世界(2019年10期)2019-11-25 07:46:04
现代园艺(2018年2期)2018-03-15 08:00:04
天津诗人(2017年2期)2017-11-29 01:24:16
新课程·下旬(2017年7期)2017-08-14 11:12:50
中国船检(2017年3期)2017-05-18 11:43:21
新农业(2016年23期)2016-08-16 03:38:14
华东经济管理(2015年6期)2015-11-14 00:07:10
西北林学院学报(2015年5期)2015-01-03 05:12:20
