
川西亚高山不同森林类型土壤呼吸和总硝化速率的季节动态
2019-02-20 10:43:04杨洪国史作民刘千里
生态学报 2019年2期
刘 顺,杨洪国,罗 达,史作民,2,*,刘千里,张 利
1 中国林业科学研究院森林生态环境与保护研究所,国家林业局森林生态环境重点实验室,北京 100091 2 南京林业大学南方现代林业协同创新中心,南京 210037 3 阿坝州林业科学技术研究所,汶川 623000
森林土壤是陆地生态系统中重要的碳氮贮存库。土壤呼吸是土壤碳输出的主要形式[1],对全球碳平衡具有重要的影响。土壤氮转化是调控森林生产力、土壤肥力等生态系统服务的重要生态过程[2]。土壤碳氮过程受多种因素的影响,在较大的空间尺度,土壤碳氮转化过程主要受气候条件和基质质量的影响[3];而在较小的尺度下,主要受植被类型、地形、土壤性状和经营干扰[4- 6]等的影响。林分类型的改变将对土壤碳氮库及其生物化学过程产生重要的影响[7],主要是受土壤凋落物的输入、土壤微生物及水热状况等因素的调控[8- 10]。但也有研究表明土壤碳氮矿化并不受林型的影响[11]。尽管土壤碳氮转化过程已经开展了大量的研究,但是特定的研究多偏重于其中一方面。有研究指出土壤碳氮循环存在耦合性[12],土壤碳氮矿化的异质程度是调控土壤有效氮供应、植物生产力和影响陆地生态系统碳收支的重要因素[13]。
川西亚高山林区位于青藏高原东南缘,地形复杂、生态脆弱,是长江上游重要的水源涵养地和生态屏障。该区域原有的主要森林类型是以岷江冷杉为优势种的亚高山暗针叶林,但其在20世纪50—80年代,经历了大规模的砍伐利用,随后进行了以天然、人工、天然和人工共同促进的更新方式,从而形成了多样的森林类型。对于该区域森林土壤碳氮转化过程的研究多偏重于土壤碳矿化过程(土壤呼吸速率)的研究[14- 16],但得出的结果并不一致。Chen等[16]的研究结果显示原始林土壤呼吸速率大于次生林,土壤呼吸与土壤温度和水分显著相关。但Luo等[15, 17]研究发现原始林土壤呼吸速率小于次生林,土壤呼吸主要受土壤温度的调控,土壤水分对其影响不大。而关于该区域森林土壤氮转化过程的研究多见于净矿化速率的研究[18- 20]。原始林与其砍伐后通过不同恢复措施形成的不同森林类型在凋落物输入和土壤环境等方面发生了改变,进而会影响土壤呼吸和总硝化速率。目前,关于该区域不同森林类型土壤呼吸和总硝化速率对比分析的研究较少[21],不同森林类型土壤呼吸和总硝化速率的影响因素及土壤呼吸和总硝化速率是否存在耦合关系仍不清楚。
气压过程分离系统(Barometric Process Separation System,BaPS)是测定土壤呼吸速率和总硝化速率的仪器,具有操作简单,经济适用的优点,已被用于土壤呼吸速率和总硝化速率的测定中[9, 22]。因此,本文采用BaPS技术对川西理县毕棚沟4种不同类型森林(岷江冷杉原始林、粗枝云杉阔叶林、红桦-岷江冷杉天然次生林和粗枝云杉人工林)土壤呼吸和总硝化速率进行了测定,目的在于(1)评估不同林型土壤呼吸和总硝化速率季节动态特征;(2)分析土壤呼吸和总硝化速率的影响因素;(3)描述土壤呼吸和总硝化速率间的关系,旨在为区域生态恢复建设和调控管理提供科学依据。
1 研究区概况
研究地点位于四川省阿坝藏族羌族自治州理县境内的毕棚沟自然保护区(31°14′—31°19′ N,102°53′—102°57′ E),海拔2458—4619 m,属于青藏高原-四川盆地的过渡地带。年均气温2.7℃,最高气温23℃(7月),最低气温-18℃(1月);年均降水量850 mm,多集中在生长季。生长季一般为5月到10月,冬季气温较低,存在季节性雪被覆盖期。研究区内原有主要森林类型为亚高山原始暗针叶林[23]。在20世纪50—80年代,以冷杉为优势树种的原始暗针叶林被大面积的采伐利用,之后进行了以云杉为主的人工更新,同时,在一部分地区进行着以桦木等阔叶先锋树种为主的天然更新[24]。1998年,天然林资源保护工程启动,该区森林禁伐封育,现已形成人工林、天然次生林以及人工、天然更新共同作用形成的林分镶嵌分布[25]。典型优势乔木有岷江冷杉(Abiesfaxoniana)、红桦(Betulaalbo-sinensis)和粗枝云杉(Piceaasperata)等;灌木主要有康定柳(Salixparaplesia)、箭竹(Fargesiaspathacea)、红毛花楸(Sorbusrufopilosa)、高山杜鹃(Rhododendrondelavayi)和三颗针(Berberisjulianae)等;主要草本为苔草(Carextristachya)、蟹甲草(Cacaliaauriculata)和高山冷蕨(Cystopterismontana)等。土壤为山地棕壤,石砾含量较多。
2 研究方法
2.1 样地设置
本研究选择4种类型森林作为对象,分别为岷江冷杉原始林、粗枝云杉人工林、粗枝云杉阔叶林(其中粗枝云杉为人工栽植,后无人为经营管理,林内混生红桦等阔叶树)和红桦-岷江冷杉天然次生林。岷江冷杉林海拔约3500 m,林龄约192年;3种恢复林分海拔在3000 m左右,林龄约30年。岷江冷杉原始林和粗枝云杉人工林采用随机设置的方法;由于采伐后粗枝云杉的栽植方式为块状,所以形成了粗枝云杉阔叶林和红桦-岷江冷杉天然次生林交替分布的格局,选择交替分布的粗枝云杉阔叶林和红桦-岷江冷杉天然次生林进行样地布设,各森林类型均设置3块20 m×20 m的样地作为重复。各林分的具体情况见表1。

表1 不同森林类型的基本情况
*pH测定时水土比为1:2.5;“—”表示林地较平缓,未记坡度和坡向;SE:东南坡向,southeast slope;SOC:土壤有机碳,soil organic carbon;TN:全氮,total nitrogen;C/N:碳氮比,the ratio of SOC to TN;SMBC:土壤微生物量碳,soil microbial biomass carbon;SMBN:土壤微生物量氮,soil microbial biomass nitrogen;FRB:活细根(Ø≤2 mm)生物量,living fine root biomass
2.2 土壤呼吸和总硝化速率
从2015年5月到11月间,每月进行一次土样采集,供土壤呼吸和总硝化速率测定。样品采集采用环刀(容积为100 cm3)原位取样,在BaPS系统中,以5个环刀土壤样品为一组,5个环刀土样均为在同一个样地内按“S”型采集的土壤。采样时,先用小刀轻轻地清除地表凋落物和动植物残体,然后切入环刀,用锤子轻轻敲打环刀边缘使其刚好没入土中。之后,小心取出环刀,用小刀将环刀下部土壤削平,并迅速盖好盖子,防止水分散失。采样结束后,立即将装有土样的环刀带回实验室置于4 ℃冰箱中保存,并于一周内完成测定。

2.3 土壤温度和水分
在每次采样的同时,采用便携式数字温度计记录采样点5 cm处的温度。BaPS测定结束后,将5个环刀土样取出,105 ℃烘箱中烘干测定土壤含水量和容重。考虑到土壤质地的影响,数据分析中水分指标是土壤呼吸和总硝化作用密切相关的土壤充水孔隙度(Water-filled pore space,WFPS),它代表了土壤的水分饱和度,计算公式如下:
WFPS=(SWC ×BD)/STP
STP=(1 -BD/PD) ×100%
式中,WFPS为土壤充水孔隙度,%;SWC为土壤重量含水量,%;BD为土壤容重,g/cm3;STP为土壤总孔隙度,%;PD为土壤颗粒密度,取2.65 g/cm3。
土壤温度和水分的季节变化如图1所示。
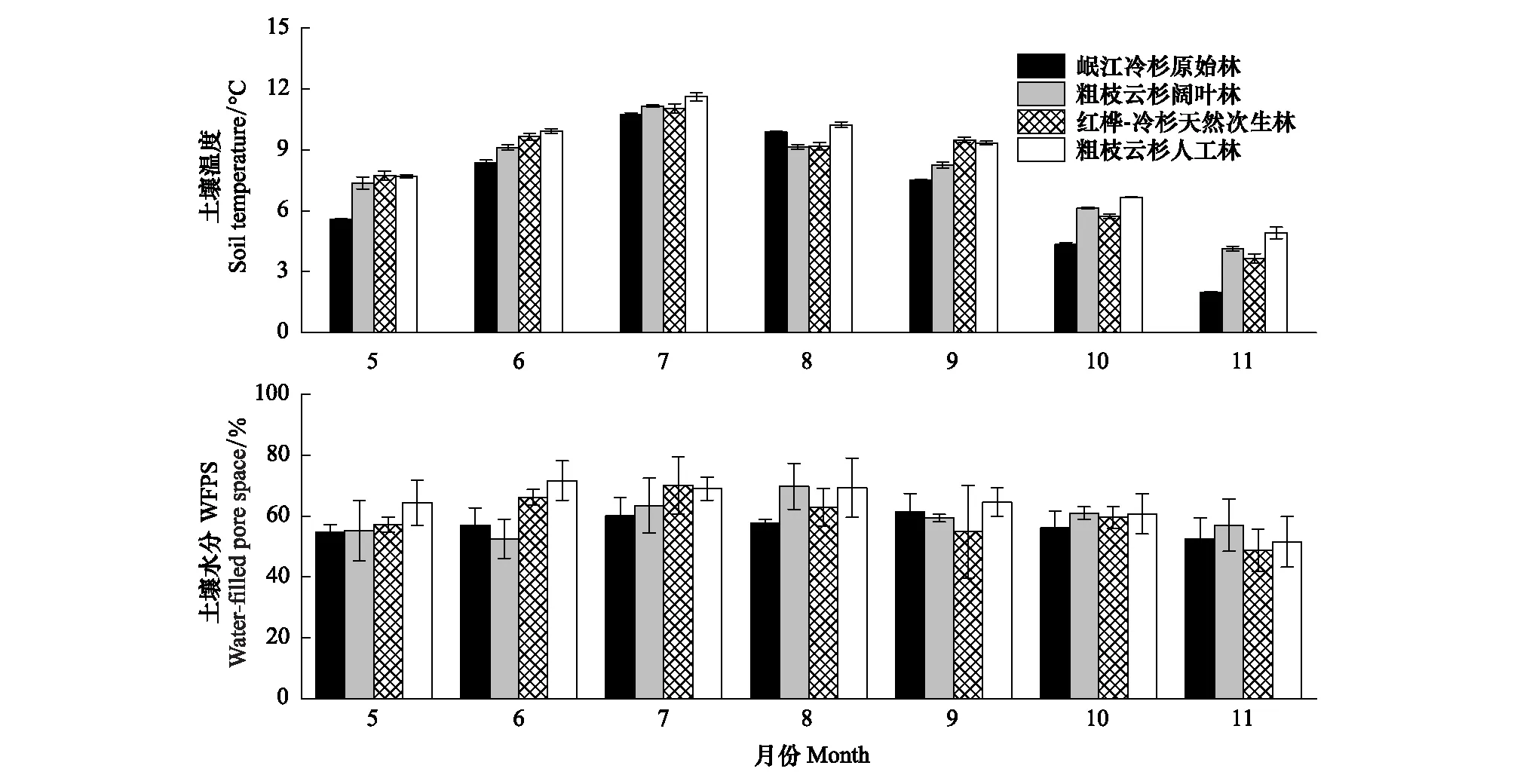
图1 不同林分土壤温度和水分的季节变化Fig.1 Seasonal dynamics of soil temperature and soil water filled pore space (WFPS) in different forests
2.4 数据处理
所有变量在分析前进行异质性检验(Levene′s test)。月份、林型对土壤呼吸和总硝化速率的影响采用双因素方差分析(two-way ANOVA)。所有的检验显著性水平为α=0.05,多重比较采用Tukey HSD法。土壤呼吸速率与土壤温度间的关系采用指数方程拟合:y=aebx(y为土壤呼吸速率,x为土壤温度,a和b为系数);土壤呼吸的温度敏感性Q10计算如下:Q10=e10b(b为土壤呼吸速率与温度指数关系中的系数)。土壤总硝化速率与土壤温度间的关系采用线性方程拟合。以土壤呼吸和硝化速率为因变量,以细根生物量、凋落物量、土壤pH、有机质、C/N、微生物量碳和微生物量氮为自变量,分别进行逐步回归分析。数据的处理和分析采用SPSS软件进行,图片采用Origin 8.0绘制。
3 结果与分析
3.1 土壤呼吸和总硝化速率

图2 不同林分土壤呼吸和总硝化速率的季节变化Fig.2 Seasonal dynamics of soil respiration and gross nitrification rate in different forests不同小写字母表示同一林型不同月份间差异显著,不同大写字母表示相同月份不同林型间差异显著,无对应字母表示差异不显著

指标Indices林型 Forest type月份 Month交互作用 InteractiondfFdfFdfF土壤呼吸速率Soil respiration rate325.617***626.850***181.059NS土壤总硝化速率Soil gross nitrification rate336.160***652.891***187.758***
***P<0.001,NS:不显著,non-significant
3.2 土壤呼吸和总硝化速率与环境因子的关系
对土壤呼吸和总硝化速率与土壤温度和水分进行拟合分析,可知土壤呼吸和土壤温度间呈指数相关的趋势,土壤总硝化速率与土壤温度呈线性相关的趋势;土壤呼吸和总硝化速率与土壤水分间的相关性不显著(图3)。表明水分不是影响本研究区森林土壤呼吸和总硝化速率的限制因素。通过土壤呼吸速率与土壤温度的指数方程系数可以计算出土壤呼吸Q10值(表3)。不同森林土壤Q10值以岷江冷杉原始林最高,为4.71,而以粗枝云杉人工林最小为2.59。岷江冷杉原始林Q10较高的原因可能与其海拔较高有关。

图3 不同林分土壤呼吸、总硝化速率与温度和水分间的关系Fig.3 Relationships between soil respiration, gross nitrification rate and soil temperature and WFPS

林分 Forest types方程 ModelsR2PQ10土壤呼吸-温度Soil respiration rate-Temperature岷江冷杉原始林 Abies faxoniana primary foresty = 44.63e0.155 x0.888<0.0014.71粗枝云杉阔叶林 Picea asperata broadleaved mixed foresty = 117.96e0.127 x0.610<0.0013.56红桦-岷江冷杉天然次生林 Natural secondary Betula-Abies for-esty = 85.89e0.112 x0.2530.0203.06粗枝云杉人工林 Picea asperata plantation foresty = 140.27e0.095 x0.3720.0032.59土壤总硝化-温度Soil gross nitrification rate-Temperature岷江冷杉原始林 Abies faxoniana primary foresty = 5.86 x+3.050.651<0.001粗枝云杉阔叶林 Picea asperata broadleaved mixed foresty = 5.52 x-4.810.667<0.001红桦-岷江冷杉天然次生林 Natural secondary Betula-Abies for-esty = 3.89 x+1.530.3710.003粗枝云杉人工林 Picea asperata plantation foresty = 5.26 x-6.230.634<0.001
3.3 土壤呼吸和总硝化速率在林型间变异的影响因子
对不同林型土壤呼吸和总硝化速率与其相关的影响因子进行逐步回归分析(表4),结果表明在不同林型间,凋落物量、土壤pH和有机质含量对土壤呼吸和总硝化速率具有显著影响。三者分别解释了土壤呼吸和总硝化速率变异的92.9%和89.7%。凋落物量与土壤呼吸和总硝化速率的回归系数均为负值,表明凋落物的量对土壤呼吸和总硝化速率有一定的抑制作用,土壤pH和有机质均对土壤呼吸和总硝化速率有促进作用。

表4 土壤呼吸和总硝化速率与相关因子的逐步回归方程
n=12;LM为凋落物量Litter mass;SOM为土壤有机质Soil organic matter
3.4 土壤呼吸和总硝化速率的关系
对不同林分土壤呼吸速率和总硝化速率进行线性拟合(图4),得出不同林分土壤呼吸速率和总硝化速率间均存在着极显著的正相关性(P<0.01),决定系数R2介于0.330—0.774之间。粗枝云杉阔叶林和红桦-岷江冷杉天然次生林土壤总硝化速率随着土壤呼吸增加的增长趋势相对比较平缓,线性拟合方程斜率分别为0.066和0.082,而其他2种林分拟合方程的斜率均大于0.1。

图4 不同林分土壤呼吸和总硝化速率间的关系Fig.4 Relationships between soil respiration and gross nitrification rate in different forests
4 讨论
土壤呼吸和总硝化速率受许多因素的影响,如微生物、基质和环境状况等[29-30]。这些因素会受到林型的影响,从而可能会造成不同林型间土壤呼吸和总硝化速率等的差异[8, 11]。本研究中不同林型间凋落物量、土壤pH和有机质含量对土壤呼吸和总硝化速率有显著影响。土壤有机质和pH促进了土壤呼吸和总硝化速率。土壤有机质是有机物分解过程的底物,是微生物能量获取的来源,因此较高的有机质可以支撑土壤中较高的微生物活性,促进土壤呼吸和硝化作用[31]。土壤pH正效应可能有以下两个原因:第一,在一定程度上,提高土壤pH,可以增加基质的有效性[32];第二,较低的pH会对土壤微生物造成毒害作用,抑制微生物活性[33],进而降低土壤呼吸和总硝化速率。凋落物被认为是土壤有机质的重要来源,在一定程度上可以反映基质的有效性[2]。但本研究得出凋落物量对土壤呼吸和总硝化速率具有抑制作用,可能是与本研究区年均气温较低,凋落物分解速率较慢,凋落物层较厚,土壤中CO2向大气的扩散受阻[34],降低了土壤氧有效性,不利于微生物活动[35],并有可能改变了土壤微生物群落结构。土壤呼吸和总硝化速率是微生物调控的过程,而土壤微生物量碳氮并未对土壤呼吸和总硝化速率产生显著影响,但以土壤呼吸和总硝化速率较高的粗枝云杉阔叶林较高(图2和表1)。另外,粗枝云杉人工林较高的细根生物量(表1),也可能是导致其土壤呼吸速率较高的原因。
森林的经营管理方式等人为干扰会对生态系统功能产生长期的影响。有研究表明针阔混交林更有利于土壤团聚体的稳定[36],并且在森林恢复过程中,红桦-岷江冷杉天然次生林土壤受到人为扰动的影响较小,对土壤团聚体的破坏性较小,使得团聚体中有机质得到一定程度的保护,会减缓土壤有机质的矿化等[37],这也可能是造成红桦-岷江冷杉天然次生林土壤呼吸和总硝化速率略小于其他两种恢复林分的原因。岷江冷杉原始林土壤呼吸和总硝化速率较低与其所处海拔最高,年均气温较低有关。
土壤温度和水分一般被认为是引起土壤呼吸和总硝化速率季节动态的主要影响因子[22, 38]。本研究中4种林型土壤呼吸速率和总硝化速率具有明显的季节变化,表现为生长季初期和末期较低,而中期速率较高的单峰变化趋势,与多数研究结果一致[27]。土壤呼吸和总硝化速率从5—7月逐渐升高,以7月份达到最高,之后逐渐下降(图2),与土壤温度和水分在季节间的变化趋势总体一致(图1)。土壤呼吸与土壤温度呈显著指数相关,而与土壤水分相关性不显著(图3),这与川西亚高山原始林[17]、天然和人工云杉林[39]等研究结果一致。与土壤呼吸相似,土壤总硝化速率与土壤温度显著正相关,但与土壤水分相关性不显著(图3),与刘义等[40]研究得出的土壤总硝化速率与土壤水分显著正相关的结果不一致,这可能与本研究对象土壤水分的季节变异较小有关。土壤温度主要是通过影响根系活力、有机质分解和微生物活性等改变土壤碳氮转化速率[30, 41]。土壤水分则是通过影响根和微生物活动、底物的扩散和氧气浓度而直接和间接的影响土壤碳氮转化速率[42-43]。在土壤水分不是环境限制因子时,土壤温度通常是影响土壤呼吸的重要影响因子[44]。但当土壤水分低于某一阈值时,土壤碳氮转化速率受到溶质扩散和微生物活性的制约,一般和土壤水分呈正相关关系;但是土壤水分超过某一阈值,土壤氧气供应不足,则会限制土壤碳氮转化速率[43, 45]。本研究中,土壤水分不是本地区的限制因子[46],且在整个生长季波动变化较温度平缓,其对土壤呼吸季节动态的影响被季节波动变化较大的土壤温度所掩盖[47]。
土壤呼吸的温度敏感性(Q10)在区域和全球尺度上都具有较高的变异性,植被类型是其重要的影响因素[48]。本研究得出川西亚高山4种林型土壤Q10(表3)介于我国森林土壤Q10变化范围(1.10—5.18)之内[49]。有研究表明土壤温度波动范围越小,Q10越大[50],同时年均温度每升高1 ℃,Q10下降3.8%—8%[49,51-52]。不同林型间以岷江冷杉原始林Q10最高,可能是由于岷江冷杉原始林海拔较高,年均温度较低引起。这与Xu等[49]得出的我国森林土壤Q10与海拔正相关的结果一致。岷江冷杉原始林土壤呼吸较高的温度敏感性,也表明了高海拔和温度较低的区域可能更易受到全球气温升高的影响[53]。不同恢复林分类型的土壤Q10值有差异,以粗枝云杉阔叶林土壤Q10值最大,粗枝云杉人工林最小。Zheng等[54]对中国不同林型森林土壤Q10研究得出落叶林土壤Q10显著高于常绿林;Curiel等[55]也得出相同气候和土壤条件下的落叶林土壤Q10显著高于针叶林,主要由落叶林植物活性和物候较常绿林更具波动性引起。本研究中粗枝云杉阔叶林和红桦-岷江冷杉天然次生林中有较高比例的落叶树种,而粗枝云杉人工林为常绿针叶纯林,物种特性可能是导致3种林型间Q10变化的重要原因。另外,林型对土壤微生物群落和有机碳组分的影响也是导致Q10在不同林型间差异的重要因素[54]。
土壤氮的矿化过程是微生物利用可利用的碳源分解底物获取氮的过程,其中一部分氮被微生物固持,另外的被释放到土壤中[56]。因此大多数研究得出土壤碳氮耦合关系为正相关[57]。本研究也得出川西亚高山4种林型土壤呼吸速率和总硝化速率呈正相关关系(图4)。而当土壤碳氮含量或其比值和微生物利用效率等发生变化时,土壤碳氮转化速率的相关关系可能会发生明显的变化[58]。由于土壤中碳氮转化过程是一个微生物参与调控的过程,因此后续研究中将加强微生物在调控碳氮转化过程及其耦合关系中的作用研究。
猜你喜欢
江苏农业学报(2022年1期)2022-03-16 02:38:57
山西林业科技(2022年4期)2022-03-12 04:39:48
散文诗世界(2021年12期)2021-12-17 19:19:59
西北林学院学报(2021年6期)2021-12-07 03:26:46
岷峨诗稿(2019年4期)2019-04-20 09:02:06
花火A(2019年1期)2019-03-22 02:27:42
东北农业大学学报(2018年11期)2018-12-21 03:48:38
阿来研究(2018年1期)2018-07-10 09:39:08
现代养生·上半月(2017年7期)2017-07-07 21:43:25
防护林科技(2017年4期)2017-04-27 08:51:38
