
川西亚高山五种主要森林类型凋落物组成及动态
2019-02-20 10:43:02张远东刘彦春顾峰雪郭明明刘世荣
生态学报 2019年2期
张远东,刘彦春,顾峰雪,郭明明,缪 宁,刘世荣,
1 中国林业科学研究院森林生态环境与保护研究所,国家林业局森林生态环境重点实验室,北京 100091 2 河南大学生命科学学院,开封 475004 3 中国农业科学院农业环境与可持续发展研究所,农业部旱作节水农业重点实验室,北京 100081 4 国际竹藤中心,北京 100102 5 四川大学生命科学学院,生物资源与生态环境教育部重点实验室,成都 610064
森林冠层凋落是营养元素由地上植被转移至土壤的主要生物学途径[1-2]。凋落物作为养分的基本载体,是森林土壤有机质补充与输入的主要来源,在调节生态系统能量流动、营养物质循环及改善森林土壤质量方面扮演重要角色[3],不同森林类型凋落物产量的季节动态及凋落速率差异是影响系统内部养分周转与植物个体生长发育的重要因素,并影响着土壤碳库的大小。因此,凋落物的动态变化是森林生态系统研究不可或缺的重要过程之一,森林凋落物研究很早就受到国内外学者的广泛关注[1-3]。不同气候区森林生态系统因物种组成、群落结构等的差异导致凋落量存在较大的变异性[4]。Meentemeyer等[5]在1982年估算全球陆地植被总凋落量为54.8×109t,叶凋落量为35.1×109t,全球森林生态系统年均凋落产量为1.6—9.2 t hm-2a-1[6]。同一地区不同恢复途径下形成的森林植被凋落物产量也存在较大差异。Pandey等[7]研究印度东北部亚热带栎树林,发现天然林凋落量(547.7 g/m2)高于人工林(419.9 g/m2),天然林营养元素的周转效率要高于人工林。
川西亚高山森林是维护长江上游生态安全的绿色屏障。20世纪60—80年代,该地区老龄暗针叶林经历了大规模集中采伐,目前恢复形成多种森林类型的镶嵌分布格局[8]。不同植被类型的生态效益存在差异,对维持地区整体性生态服务功能贡献不同。近年来学者已就不同森林类型的水文效应[9],群落结构与物种多样性[10-11]、生物量与生产力[12-13]、碳储量[14]、土壤性状[15-16]等方面开展众多研究。凋落物作为碳循环中联系地上与地下碳库的纽带,对探讨土壤微生物群落、土壤理化性质、土壤碳储量等问题有重要意义,然而该区不同森林类型的凋落产量动态研究尚不够全面,限制了对不同恢复途径下森林碳循环规律的评价[2, 15, 17-19]。因此,本文以川西亚高山林区残留老龄林(VF)、次生桦木林(BF)、次生针阔混交林(MF)、人工云杉林(AF)、高山栎灌丛(AO)五种典型森林植被为研究对象,分析了各森林类型在不同生长季时期的凋落物组成及动态,为进一步揭示亚高山林区不同恢复途径下的碳循环规律提供基础数据与科学依据。
1 材料与方法
1.1 研究区概况
研究区位于四川省阿坝州理县米亚罗林区,地理坐标31°24′—31°55′N,102°35′—103°4′E,海拔2700—4200 m。该区属于青藏高原东南缘,具有典型的高山峡谷地貌。气候受着高原地形的决定性影响,属冬寒夏凉的高山气候。年平均气温6—12℃,1月份平均气温-8℃,7月份平均气温12.6℃,年降水量为600—1100 mm,森林土壤类型主要为山地棕壤。该地区典型森林植被为亚高山暗针叶林。主要乔木树种为岷江冷杉(Abiesfaxoniana)、粗枝云杉(Piceaasperata)、紫果云杉(Piceapurpurea)、红桦(Betuaalbo-sinensis),灌木树种有川滇高山栎(Quercusaquifolioides)、红刺悬钩子(Rubusaurantiacus)、野樱桃(Prunstatsienensis)、红脉忍冬(Loniceranervosa)、紫花卫矛(Euonymusporphyreus)和多种荚蒾(Viburnumspp.)等。米亚罗林区植被类型与生境多样,因海拔及坡向而分异。经历近50年来自然恢复与人工更新[20],逐渐形成残留老龄林(VF,20世纪60—80年代大规模采伐后,由于交通不便或保留母树林的需要而残留下的云冷杉暗针叶林)、次生桦木林(BF,采伐迹地上自然恢复形成的桦木纯林)、次生针阔混交林(MF,采伐迹地上自然恢复的桦木冷杉混交林)、人工云杉林(AF,粗枝云杉林)、高山栎灌丛(AO,川滇高山栎)等森林植被类型镶嵌分布的景观格局[8]。
1.2 研究方法
本研究样地选于川西米亚罗林区五种典型森林植被,分别在米亚罗镇对坡、夹壁沟左右两坡、283营林站设置样地,各类型3个样地选于不同地点。残留老龄林样地大小50 m×50 m,其余四种森林类型的样地大小均为20 m×20 m。除川滇高山栎灌丛布设2个样地外,其他森林类型各设置3个重复样地,共14个样地,基本情况见表1。实测样地内每棵树木的树高、胸径,选择与平均胸径和平均树高相近且长势健康的林木2—3株,计数年轮求得林龄。老龄林林龄在200年以上,其他四种类型林龄在30—40年,属中龄林。各样地海拔、坡度、坡向基本一致,无明显人为、牲畜干扰。
每个样地随机布置5个0.5 m×0.5 m用1 mm尼龙网制成的凋落物收集框,距地面高度0.5 m。由于该区受高山气候影响,生长季较短,集中于5—10月,而生长前期相对较长。因此在全年尺度对凋落物收集3次,依次定义为生长前期 (前一年10月1日至当年4月30日)、生长旺盛期(当年5月1日至7月31日)、生长季后期(当年8月1日至9月30日)。各收集框凋落物按叶片、枝、皮、花及果实4种器官分类,并于80℃下烘干12小时,称重并计算样地单位面积凋落物产量。空间变异性采用变异系数CV (Coefficient of variation)来描述。
1.3 数据处理
数据分析在SPSS 13.0软件下完成,采用Kolmogorov-Smirnova对数据分布进行正态性检验基础上,使用one-way ANOVA进行单因素方差分析与多重比较。绘图在Sigma Plot 10.0软件中完成。
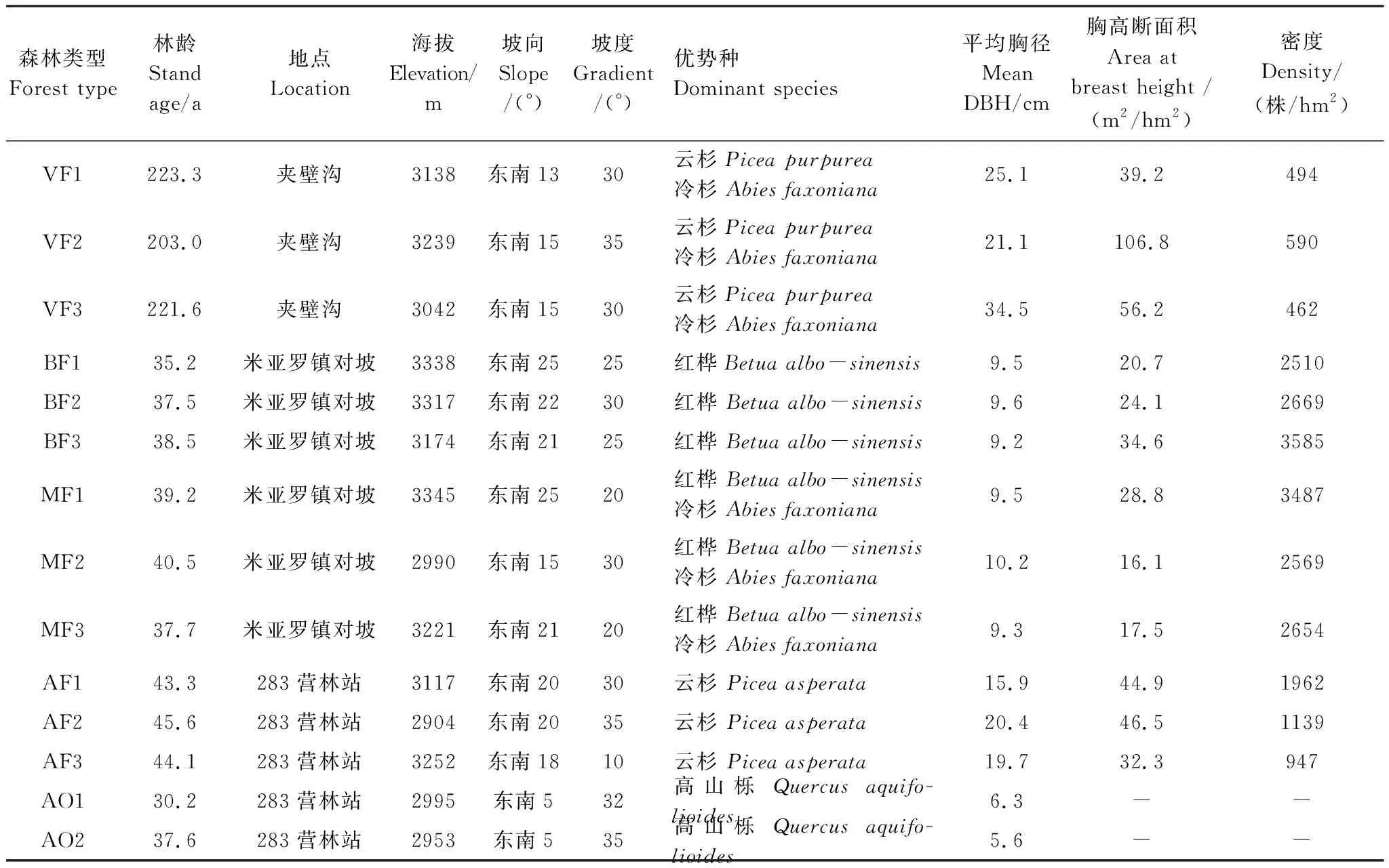
表1 研究样地基本概况
VF: 老龄林, Virgin forest; BF:次生桦木林, Secondary birch forest; MF: 次生针阔混交林, Secondary coniferous and broad-leaved mixed forest; AF:人工云杉林, Artificial spruce forest; AO: 高山栎林Alpine oak; 高山栎灌丛繁密, 未做每木检尺, 密度与胸高断面积未知
2 结果与分析
2.1 各森林类型的凋落物器官在不同时间的分配差异
树种组成及群落结构的差异会导致不同森林类型的凋落物产量存在差异。老龄林、人工云杉林、高山栎灌丛均在生长前期达到最大凋落量,分别为2.41,1.29,1.63 t/hm2(表2),占全年总凋落量的57%,52%,54%,;次生桦木林、次生针阔混交林凋落产量则在生长季后期到达最大值,分别为1.34,1.80 t/hm2,占总凋落量的51%,51%(表2)。在全部凋落期内,叶片是五种森林类型凋落物中的主要构成成分,比例均在60%以上(表2)。因此,不同森林类型的叶片凋落动态与林分总凋落产量变化表现出一致的趋势。五种森林类型的枝凋落量均在生长前期达到最大值,分别占全年枝凋落量的70%,56%,67%,72%,41%(表2)。高山栎灌丛在三个凋落期的枝凋落量较接近(0.15,0.11,0.11 t/hm2) (表2)。各类型树皮凋落量很小,最大凋落值是生长旺盛期的人工云杉林(0.13 t/hm2占该期总量的19.1%);其他类型中,老龄林在生长旺盛期树皮凋落量较高(10.4%);次生桦木林在生长前期(11.4%)。花、果实凋落最大值为生长前期的高山栎灌丛(0.23 t/hm2,占总量的14.1%);其次为生长旺盛期、后期老龄林(14.6%,10.5%)。
2.2 相同时期不同森林类型间凋落物差异
在生长前期,老龄林总凋落量、叶凋落量显著高于其他森林类型(P<0.05);次生桦木林总凋落产量显著小于其他森林类型(P<0.05);次生桦木林、高山栎林枝凋落量显著小于其他3种森林类型(P<0.05)。生长旺盛期,次生桦木林总凋落量显著高于老龄林、高山栎林(P<0.05);而后二者叶片凋落量则显著小于其他三种森林类型(P<0.05);该时期高山栎灌丛的枝凋落量显著高于其他几种森林类型(P<0.05),而其他类型间无显著差异(P>0.05)。生长季后期,次生桦木林、次生针阔混交林总凋落量、叶凋落量显著高于其他森林类型(P<0.05),而次生桦木林枝凋落量显著小于其他森林类型(P<0.05)(表2)。
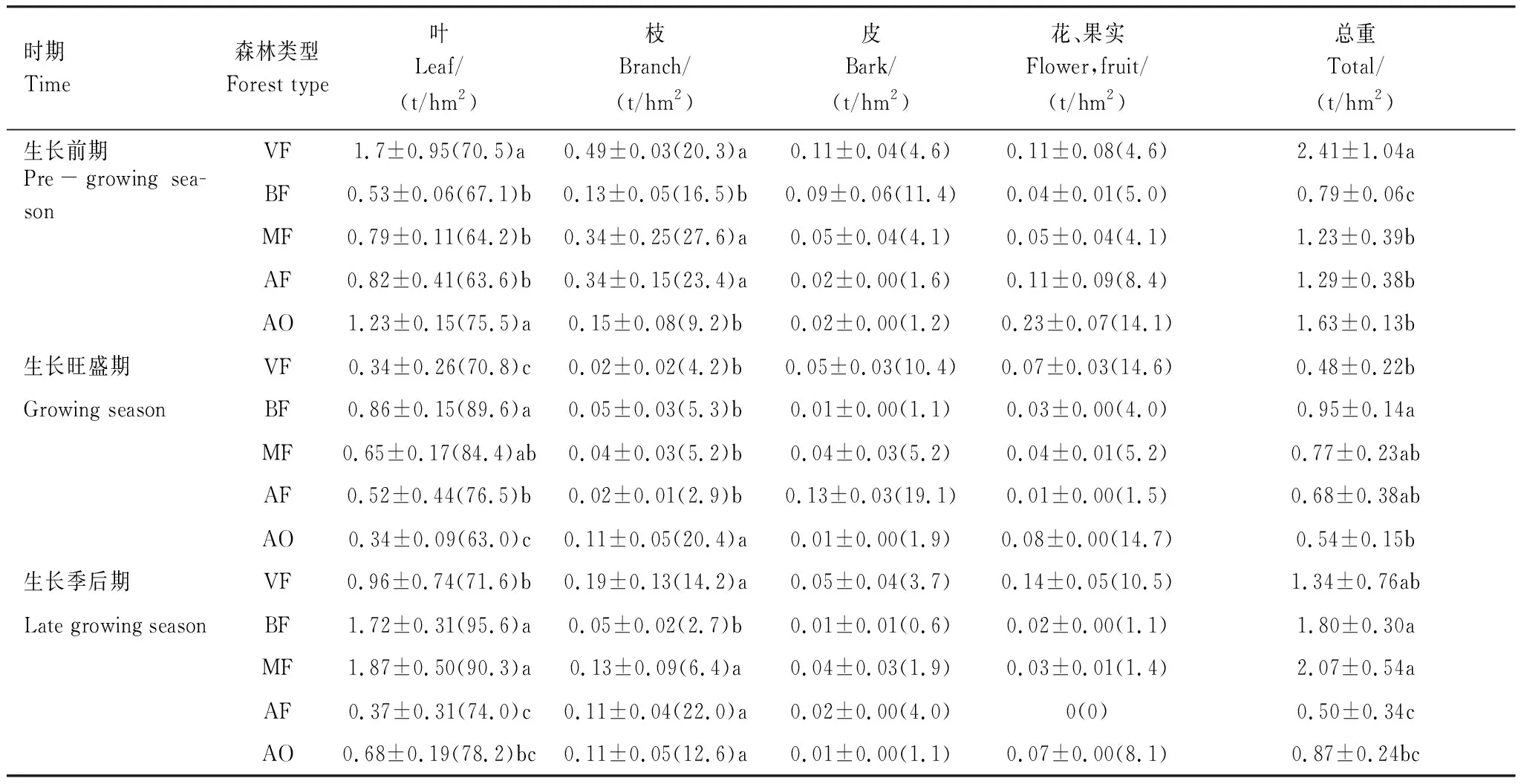
表2 五种主要森林类型的凋落物组成与动态
括弧内为各器官占总重量的百分比, 同凋落期内各列标相同小写字母者表示无显著差异,P> 0.05, 不同小写字母表示差异显著,P<0.05
2.3 不同森林类型全年凋落量与组成
由图1可知,五种森林类型的全年凋落产量大小依次为老龄林(4.32 t/hm2)、次生针阔混交林(4.10 t/hm2)、次生桦木林(3.52 t/hm2)、高山栎灌丛(3.01 t/hm2)、人工云杉林(2.34 t/hm2)。叶凋落量最大类型为次生针阔混交林(3.11 t/hm2),最小为人工云杉林(1.70 t/hm2);枝凋落量最大类型为老龄林(0.71 t/hm2),最小为次生桦木林(0.23 t/hm2);皮凋落量最大的类型为老龄林(0.21 t/hm2),最小为高山栎灌丛(0.03 t/hm2);花果实凋落量最大为高山栎灌丛(0.38 t/hm2),最小为次生桦木林(0.09 t/hm2)。4种乔木林中,人工云杉林全年凋落量显著小于其他3种森林类型(P<0.05)。高山栎灌丛全年凋落量显著小于老龄林和次生针阔混交林(P<0.01)而与另两种无显著差异(P>0.05)。各森林类型的叶片全年凋落量占总量比例均超过70%。在全年凋落量中,叶片比例最高为次生桦木林(87.8%),枝、皮比例最高为人工云杉林(18.9%,7%),花果实比例最高者为高山栎灌丛(12.7%)。
3 讨论
3.1 凋落物产量的时空变异
群落特征的时空异质性是陆地生态系统的主要表现特征。森林生态系统内部群落结构、物种组成、年龄分布的差异使生态系统表现出空间变异,同时由于季节变化,气候更替使森林生态系统表示出时间变异。生态系统的时空变异导致凋落物产量在时空分布上的差异性。如图2所示,除老龄林外,其他森林类型的样地内空间变异均高于样地间的空间变异。人工云杉林样地内空间变异最小,这主要归因于人工林相对均质的林分密度与相近的个体生长状况;次生针阔混交林样地内空间变异最大,这主要是由于其林分稀疏差异与个体分化显著所致。
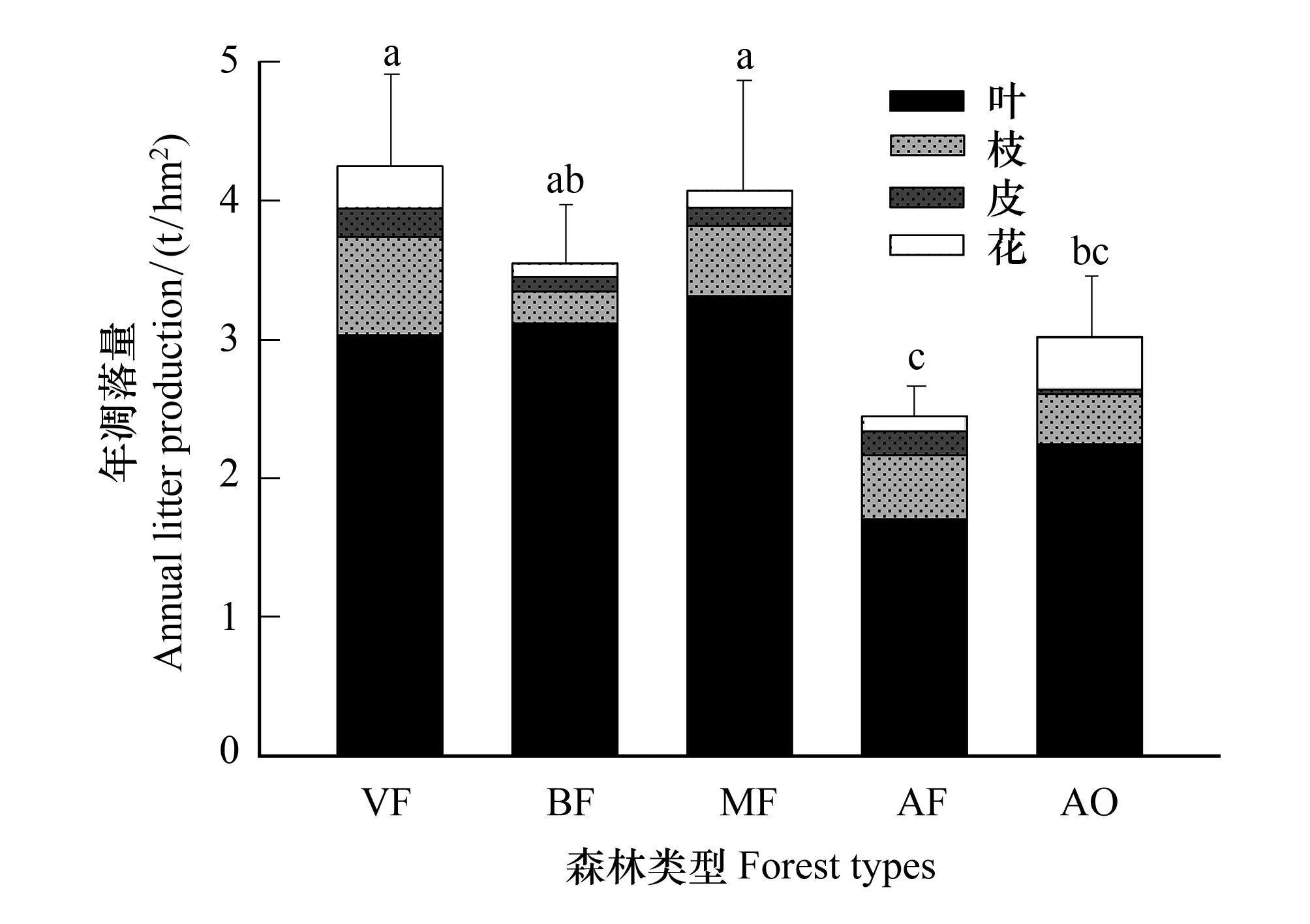
图1 五种森林类型全年凋落产量及组分构成 Fig.1 Total litter production and its composition of the whole year among five forest types不同类型间标不同字母表示在0.05水平上有显著差异(P<0.05); VF: 老龄林, Virgin forest; BF:次生桦木林, Secondary birch forest; MF: 次生针阔混交林, Secondary coniferous and broad-leaved mixed forest; AF:人工云杉林, Artificial spruce forest; AO: 高山栎林Alpine oak
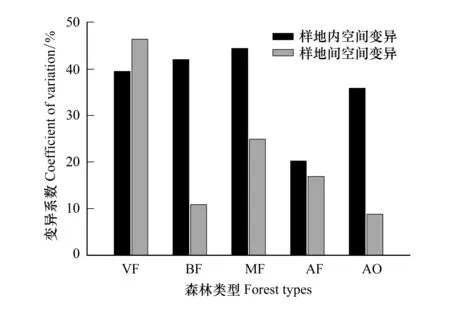
图2 不同森林类型样地内与样地间变异系数对比分析 Fig.2 Comparison of coefficient variation for each plot and among plotsCVW, 样地内空间变异,Coefficient variation within plot; CVA, 样地间空间变异, Coefficient variation among plots
通过对不同森林类型样地间凋落物空间变异分析可知(图3),对于叶片凋落物,老龄林、人工云杉林的变异系数为50%—90%,高于次生桦木林、次生针阔混交林和高山栎灌丛(10%—30%),说明林分立地条件对原始林、人工云杉林叶片凋落量的影响要高于后三者。各时期次生桦木林的变异系数均为最小,表明次生桦木林叶片凋落量对立地与环境条件差异的响应最不敏感。在时间尺度上,各类型在生长旺盛期、后期的变异系数均高于生长前期。
枝凋落物是森林生态系统中的第二大凋落组分。对于枝凋落物,次生针阔混交林的变异系数最大(73%—74%),人工云杉林最小(42%—45%)(图3);除老龄林外,其他4种类型在不同时期的变异系数近似为常数,说明在不同立地条件下相同森林类型,枝凋落物产量在3个凋落期具有相同变化趋势,即对季节变化的响应较一致。这表明,每种森林类型的枝凋落物在样地间不存在显著的差异,这可能与林地立地条件有关。
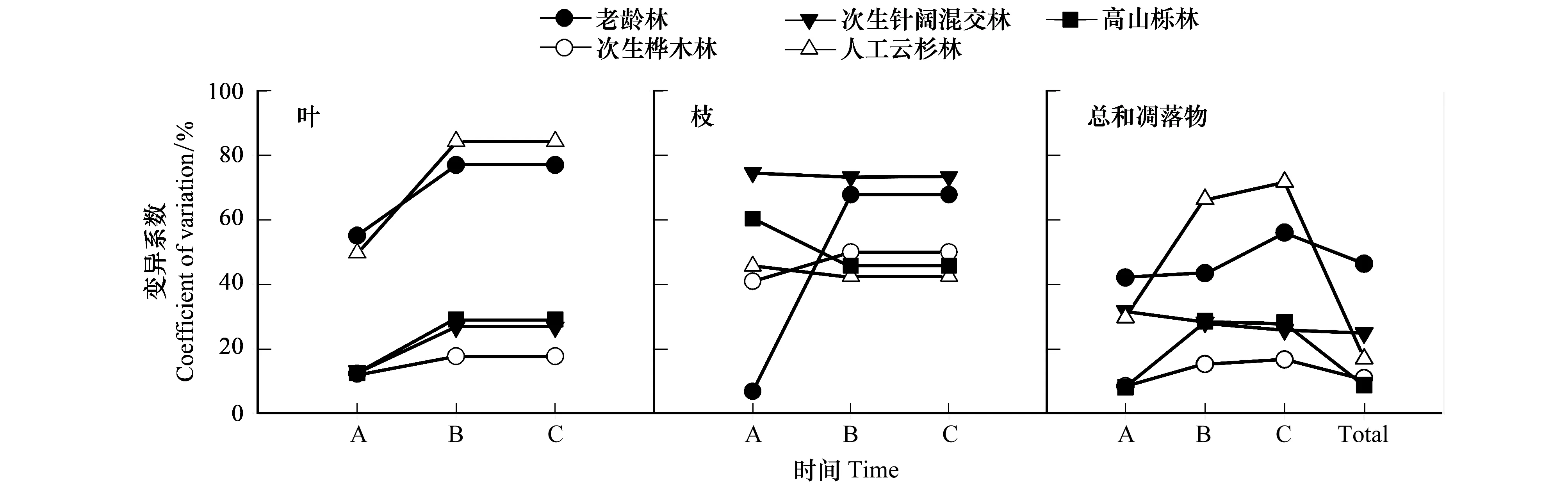
图3 主要器官与总凋落物的时空变异性Fig.3 Spatial and temporal variability of leaf, branch and total productionA, 生长前期, Pre-growing season;B, 生长旺盛期, Growing season; C,生长季后期, Late growing season; Total, 全年凋落量, Total litter production in the whole year
在总凋落物方面,全年凋落物产量样地间变异系数大小依次为:老龄林(46.4%)、次生针阔混交林(24.9%)、人工云杉林(16.9%)、次生桦木林(10.8%)、高山栎林(8.8%)。3个凋落期的次生桦木林的变异系数最小,再次表明次生桦木林对林分立地条件差异的响应较低;老龄林在生长前期的变异系数最高;人工云杉林在生长旺盛期、后期的变异系数最高,与叶片凋落特征相近,这是由于人工云杉林凋落物以叶为主(叶占69.2%),并可能与林分密度有关。
3.2 凋落物产量与林分密度、胸高断面积关系
同一气候区下,树种组成、林分密度、林分胸高断面积、林龄是决定凋落物产量的关键因子[2, 21-22]。林分密度与胸高断面积是影响凋落物大小的最直接因素,是讨论凋落物大小成因的重要指标[22]。本文运用林分密度、胸高断面积、及二者乘积对14个样地凋落量进行了线性回归分析。如表3所示,只有当林分密度、胸高断面积乘积作为自变量时,模型回归系数最高,并达到显著水平(P=0.0376)。林分密度,胸高断面积单独使用时均未达到显著水平。这是由于在林分发育过程中树木径向生长会受到林分密度的影响,当立地条件一定,资源与空间的限制作用突现时,树木径向生长与林分密度呈负相关。调查的原始林已表现出很强的自疏效应,密度很小,但胸高断面很大(表1);另外三类40年生乔木林也表现出一定强度的林内竞争,个别树木已发生干枯死亡的现象。因此使用单一林分因子表征凋落物产量是比较困难的,组合因子更能反映凋落物特征。

表3 凋落物产量及林分因子回归模型
Y=aX+b,n=12, ADH , 胸高断面积(cm2/hm2), Area at breast height;D/tree hm-2, 林分密度(株/hm2), Stand density
3.3 不同恢复途径的凋落物产量及对森林养分循环的意义
森林凋落物产量受控于生物因素(森林类型,群落结构)与非生物因素(立地条件与与气象因子)的双重影响[23]。人工更新形成的云杉林凋落物产量最小,显著低于在保留母树采伐迹地上自然恢复形成的次生针阔混交林和无保留母树迹地上自然恢复形成的次生桦木林。在自然更新模式下,针阔混交林的凋落量高于桦木林。与我国东北林区主要森林类型相比,次生针阔混交林与东北林区的混交林较一致(4.15 t/hm2),次生桦木林凋落量要略高于东北落叶阔叶林(3.13 t/hm2)。有研究表明,人工云杉林凋落物年腐解率小于阔叶林与原始林[15],即凋落物分解周期较长[18],因此相对于桦木阔叶林,其凋落物对土壤有机质和土壤养分的补充较慢。同时,人工造林往往伴随整地,扰动加速土壤碳释放,因此人工云杉林土壤易发生土壤养分缺乏与碳流失,不利于土壤碳的保存。但人工云杉林凋落物分解速度慢使得云杉林内枯落物蓄积量高于阔叶林[24],张远东等[9]研究发现,40年生人工云杉林苔藓和枯落物蓄积量显著高于同龄针阔混交林,可见人工云杉林在地被物层累积方面具有优势。本研究老龄林全年凋落量与该区Yang等[2]研究结果一致(4.32 t/hm2),略小于林波等[17]研究的结果(4.77 t/hm2),而高于贡嘎山峨眉冷杉老龄林(2.28 t/hm2)[25];人工云杉林凋落量要低于宿以明[19]和林波[17]在该区的研究结果。以上研究表明,不同恢复途径影响森林凋落物产量,人工云杉林凋落量低于次生桦木林、次生针阔混交林,并且人工云杉林对林分立地条件响应较后者敏感,因此对于川西亚高山林区而言,天然更新形成的桦木林、针阔混交林具有更普遍的适应性,较快的凋落物分解周转有利于土壤养分的及时补充,人工云杉林仅地被物累积较高。
猜你喜欢
今日中国·西班牙文版(2021年4期)2021-04-09 11:22:46
生物学通报(2020年11期)2020-10-22 01:20:20
中国水土保持科学(2019年6期)2019-04-26 05:13:48
现代园艺(2018年2期)2018-03-15 08:01:03
浙江农林大学学报(2016年3期)2016-06-30 03:08:44
广西林业科学(2016年1期)2016-03-20 05:33:02
广西林业科学(2016年4期)2016-03-16 05:44:58
甘肃林业(2016年6期)2016-02-27 11:29:12
西部林业科学(2015年6期)2016-01-27 10:39:16
吉林大学学报(医学版)(2015年1期)2015-12-17 07:47:20
