
横断山区隐瓣蝇子草四个居群的核型分析
2019-01-24 03:40:24饶培瑜胡伸萌苏富明孙文光李志敏
云南师范大学学报(自然科学版) 2019年1期
饶培瑜, 胡伸萌, 苏富明, 孙文光, 李志敏
(1.云南师范大学 生命科学学院,云南 昆明 650500;2.云南师范大学 生物能源持续开发利用教育部工程研究中心,云南 昆明 650500)
石竹科(Caryophyllaceae)蝇子草属(SileneL.)隐瓣蝇子草(Silenegonosperma)是多年生草本,主要分布于甘肃、青海、新疆、西藏、山西和河北等省区,国外中亚地区也有分布.主要分布于3 000~4 400 m海拔的高山草甸地区.蝇子草属约600种,是石竹科中数量比较多的属,而在我国约有110余种,以西北干旱草原荒漠地区和西南山区最为集中[1].在横断山区该属有49种,2变种,是蝇子草属的分布和分化中心[2].蝇子草属在中国分布的100余种中,仅有11种有细胞核型报道,而其余种的细胞学报道方面还有大量的空白.蝇子草属中的许多物种在系统学以及进化生物学中都作为模式类群被加以研究[3],因此本文对隐瓣蝇子草的细胞学进行了研究,除了对蝇子草属提供染色体的资料外,还为生物进化学等方面提供研究基础.
1 材料和方法
材料来源:隐瓣蝇子草分布于横断山高海拔地区,实验材料采自云南西北部的德钦县及四川西部的九龙县、小金县及松潘县(详见表1),凭证标本在中国科学院昆明植物研究所标本馆中(KUN)保存.
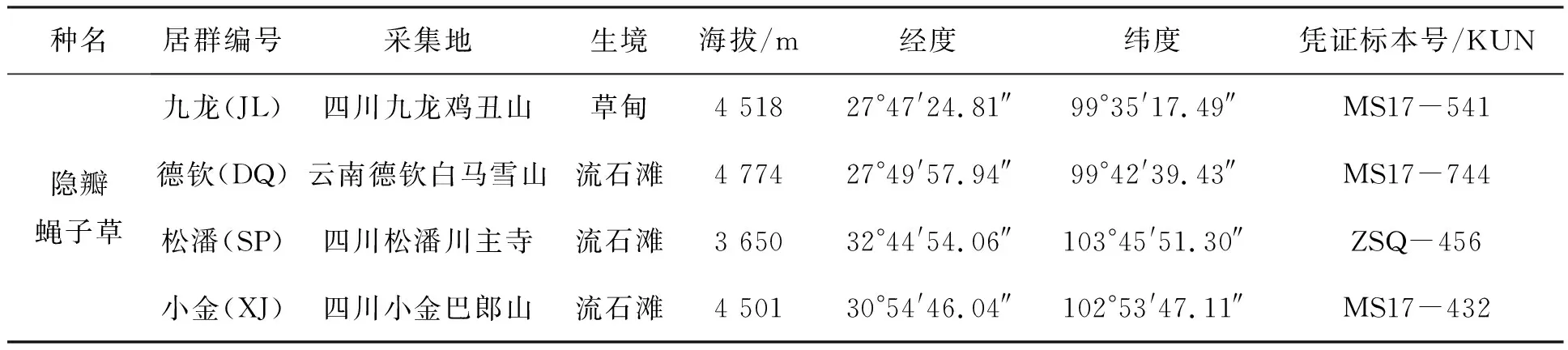
表1 横断山地区隐瓣蝇子草四个居群的来源
研究方法:本研究采用国内外通用的植物染色体常规压片法,并参考了洪德元[4]以及刘永安[5]的实验方法.首先选取饱满的种子放置在培养基中,于20 ℃的培养箱中进行萌发,并给予适当的光照;一般3~7 d左右萌发;取萌发长度为0.5~1 cm的根尖,将根尖置于浓度为0.03 mol/L的8-羟基喹啉中进行预处理,预处理时间为3 h.将预处理后的根尖用蒸馏水清洗三次后加入卡诺氏固定液(无水乙醇∶冰乙酸 = 3∶1),放进4 ℃冰箱中固定24 h;固定后的根尖用蒸馏水清洗三遍后,用1 mol/L的KCl进行低渗处理6 h.低渗处理后的根尖用蒸馏水清洗3遍,之后加入预热到60 ℃的1 mol/L的HCl,在60 ℃的恒温水浴锅中解离12 min;解离后的根尖用蒸馏水清洗三次后加入卡宝品红染液染色12 h以上;采用常规压片法进行镜检,挑取形态较好,较分散的中期细胞进行染色、观察和拍照.
计数和测量:根据李懋学的标准[6],计数细胞在30个以上,选取6个以上的测量细胞,用KaryoType-2.0对染色体进行测量与分析[7].计算染色体的长短臂比以及确定染色体类型,依据Levan的标准确定着丝粒的位置[8],按照Stebbins对称性来进行核型分析[9],根据Peruzzi等的检测方法检测核型不对称性[10-11],按Tanaka的关于间期核形态和分裂前期异染色质收缩方式的分类对四个居群的间期以及前期进行分类[12].
2 结 果
隐瓣蝇子草的染色体的数目和核型为首次报道.其四个居群的间期核形态、前期核形态以及中期细胞形态及模式图如图1所示,四个居群的中期染色体配对图如图2所示,其染色体参数如表2所示.
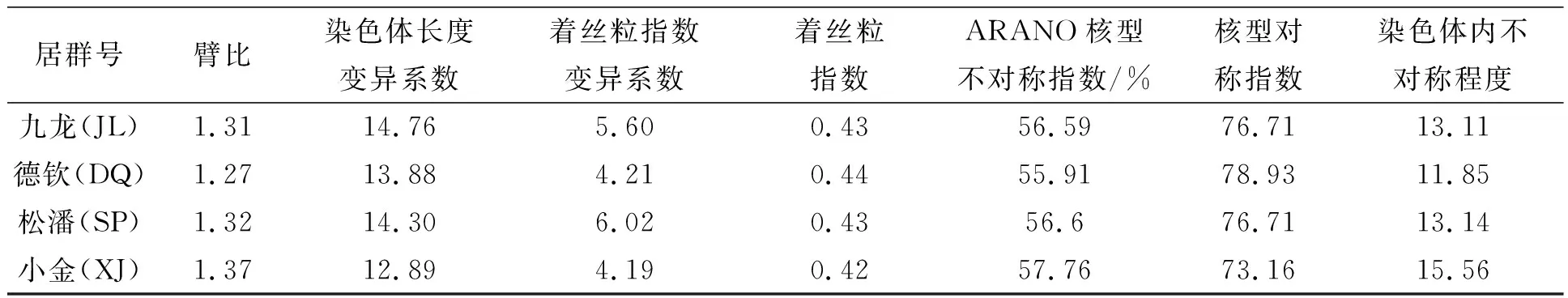
表2 隐瓣蝇子草四个居群的核型参数
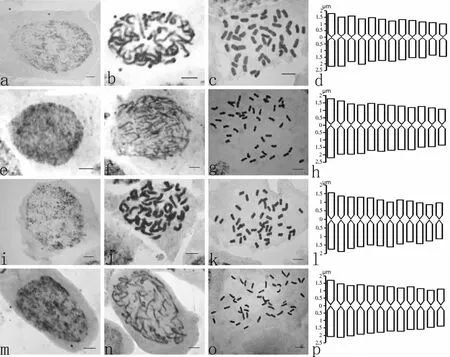
a-d:九龙居群;e-h:德钦居群;i-l:松潘居群;m-p:小金居群;标尺5 μm
Fig.1 The interphase nucleus,prophase nucleus,the chromosomes of metaphase and the mode pattern of four populations

Ⅰ:九龙居群;Ⅱ:德钦居群;Ⅲ:松潘居群;Ⅳ:小金居群;标尺5 μm
根据实验结果可知,采自四川省九龙县鸡丑山的居群为四倍体居群,此居群的间期核类型为简单染色中心型,前期核类型是中间型,核型公式为2n = 4x = 48 = 48m,核型不对称类型都为Stebbin′s-1A(见表2);云南省德钦县白马雪山顶的居群为四倍体居群,此居群的间期核类型为简单染色中心型,前期核类型是中间型,核型公式为2n = 4x = 48 = 48m,核型不对称类型都为Stebbin′s-1A(见表2);四川省松潘县主寺雪宝顶的居群的间期核类型是简单染色中心型,前期核类型为中间型,此居群为四倍体居群,核型公式为2n = 4x = 48 = 48m,核型不对称类型都为Stebbin′s-1A(见表2);小金县巴郎山居群的间期核类型是复杂染色中心型,前期核类型为中间型,为四倍体居群,核型公式为2n = 4x = 48 = 48m,核型不对称类型都为Stebbin′s-1A(见表2).四个居群的细胞学分析表明,居群间未出现染色体数目的变化,核型公式一致.
3 讨 论
隐瓣蝇子草四个居群均位于海拔3 500 m以上的高海拔流石滩地区,该区域环境较为恶劣,昼夜温差极大,紫外线辐射强烈,常年大风,造成生理性干旱[13],在这样的极端生境下,隐瓣蝇子草植株表现出一些形态适应特征如普遍低矮,根粗壮,茎、叶、花梗被短柔毛等[14],短柔毛可以反射紫外线并且减少水分的蒸发以及保温[15];本研究所采集的隐瓣蝇子草的四个居群均为四倍体,可能是通过形成多倍体来适应极端的高山环境.
从核型特征上看,这四个居群核型均属于1A型,核型变异很小,同源染色体臂比以及核型不对称系数等有一定的差异以及分化,或许暗示着居群间核型上已经产生了分化.
从染色体配对结果来看隐瓣蝇子草的四个居群都可能为同源多倍体,而同源多倍化的原因有很多,如合子的染色体加倍,分生组织的染色体加倍以及染色体未减数的配子等原因.多倍体的生境通常受到气候和地形变化影响[16],多倍体一旦由它的二倍体祖先产生,可比二倍体祖先有更好的适应性从而逐步取代二倍体个体在生境中存活下来[17].自然界中新产生并且能生存下来的同源多倍体不是本身的生存能力更强更能抢占二倍体还没有全部占领的生境,就是和其二倍体祖先的生态要求不一样从而在新的生境生存,而隐瓣蝇子草属于哪一种情况,还需要更多居群的隐瓣蝇子草的核型数据以及分析其近缘类群进一步研究后得知.
致谢:感谢中国科学院昆明植物研究所马祥光、张建文和钱栎屾在野外种子采集中的帮助.
猜你喜欢
西北植物学报(2023年6期)2023-08-14 09:24:12
农业科技通讯(2023年1期)2023-02-12 07:09:08
儿童故事画报·自然探秘(2022年12期)2022-11-24 07:29:15
今日农业(2020年14期)2020-12-14 19:47:34
诊断学(理论与实践)(2020年6期)2020-12-09 04:07:52
种子(2017年11期)2018-01-24 10:51:40
现代园艺(2017年13期)2018-01-19 02:28:17
植物研究(2017年3期)2017-11-10 10:44:12
临床与实验病理学杂志(2017年3期)2017-03-07 09:33:00
中成药(2016年4期)2016-05-17 06:07:54
