
豆粕替代鱼粉并添加精氨酸对点带石斑鱼消化酶活性和肠道组织结构的影响
2019-01-24 03:17陈思琦程镇燕安贸麟方珍珍王庆奎
中国饲料 2019年1期
陈思琦, 程镇燕, 安贸麟, 方珍珍, 王庆奎
(天津农学院水产学院,天津市水产生态及养殖重点实验室,天津300384)
点带石斑鱼(Epinephelusmalabaricus)隶属于硬骨鱼纲、鲈形目、鮨科(Serranidae),石斑鱼属其蛋白质含量高、脂肪含量低,包含多种必需氨基酸、多种无机盐和微量元素以及各种维生素,近年来,被我国东南沿海的水产养殖群体视为重要的水产品之一,但随着水产动物对鱼粉需求量的增大和全球鱼粉的产量下降,鱼粉价格直线攀升,相应的饲料成本也居高不下(叶元土等,2013),因此寻找到节约饲料鱼粉用量的适宜途径成为点带石斑鱼配合饲料开发的研究热点及难点。
目前节约饲料鱼粉用量的主要途径是寻找适宜的替代蛋白源及利用脂肪或糖类对蛋白的节约作用等方面,其中开发植物蛋白源替代鱼粉的研究则是较为活跃的部分。由于豆粕与其他植物蛋白源相比具有蛋白质含量较高、消化吸收率高、氨基酸组成较为平衡、价格合理和资源丰富等优点,一直以来都被视为理想的动物蛋白源替代品。但由于豆粕其本身含有较多抗营养因子及适口性较差,致使其在水产动物饲料中的应用受到限制。研究发现,当石斑鱼饲料中豆粕替代鱼粉比例为10%~15%时,对其生长无显著影响(罗智等,2004)。而且,大量的研究已经表明,用植物蛋白源替代鱼粉,不但会降低鱼体增重,还会影响肠道的组织结构和功能,主要表现为皱襞高度降低、顶端脱落、固有层加宽等,导致其病理学和免疫方面发生相应变化(Krogdahl等,2015;张锦秀,2003),因此,寻找如何提高植物蛋白替代鱼粉的比例而又不危害水产动物的生长及健康是亟待解决的问题。
研究表明,精氨酸及其代谢产物一氧化氮(NO)、鸟氨酸、瓜氨酸等在动物免疫防御、维持和保护肠道黏膜等方面发挥着重要的作用(Wu,2013;Gurbuz等,1998)。作为鱼类的必需氨基酸,精氨酸在鱼类营养代谢及免疫系统也发挥着重要的调节功能 (Pohlenz 等,2012;Cheng 等,2012、2011)。在对黄颡鱼及斜带石斑鱼的研究表明,饲料中适宜的精氨酸水平能有效改善鱼体生长性能,提高其肠道蛋白酶和淀粉酶的活性,增加皱襞高度和肌层厚度,对鱼体的消化产生有利影响(陈启明,2016;迟淑艳等,2016)。本试验在不同豆粕替代水平下添加精氨酸研究其对石斑鱼消化酶活力和肠道结构的影响,旨在为优化点带石斑鱼人工配合饲料提供数据支撑。
1 材料与方法
1.1 试验饲料 本试验采用3×3双因素试验设计,以鱼粉、豆粕、虾粉和谷朊粉为蛋白源,鱼油和豆油为脂肪源,其中豆粕替代鱼粉蛋白的水平分别为20%、30%、40%,精氨酸添加水平为0%、0.5%、1%,添加维生素和矿物质,并用甘氨酸来调节氮平衡,配制9种蛋白质为50%、脂肪为12%、能量为20 kJ/g的等氮等能试验饲料。记为D1(20%/0%)、D2(20%/0.5%)、D3(20%/1%)、D4(30%/0%)、D5(30%/0.5%)、D6(30%/1%)、D7(40%/0%)、D8(40%/0.5%)、D9(40%/1%)。 所有干性原料都经过粉碎机多次研磨粉碎后,过60目筛,原料逐级混匀后加入鱼油和豆油,再次混匀后加适量水,经饲料颗粒机制成直径为2mm的颗粒饲料,晾干后装入自封袋,-20℃保存备用。具体试验饲料配方及营养成分见表1,各原料及鱼体的必需氨基酸含量见表2。
1.2 试验鱼 试验鱼为天津市海发珍品实业发展有限公司提供的同一批鱼种,选取1350尾体质健壮、规格相同,个体体重(47.91±3.54)g 的点带石斑鱼,平均分为9组,每组3个重复,分别记为D1、D2、D3、D4、D5、D6、D7、D8、D9,每箱放养 50 尾,采用全封闭内循环海水养殖系统,饲养于直径为1m,高为50 cm的养殖缸中,正式试验前,试验鱼先用基础饲料驯化7 d。每天早上清除缸内粪便,并换水,每天换水量为总水量的1/3。饲养试验进行10周,期间,每天饱食投喂2次 (09:00和16:30),养殖试验期间水体温度为26~29℃,溶解氧浓度为8.0~10.0mg/L,pH为7.5~7.7。
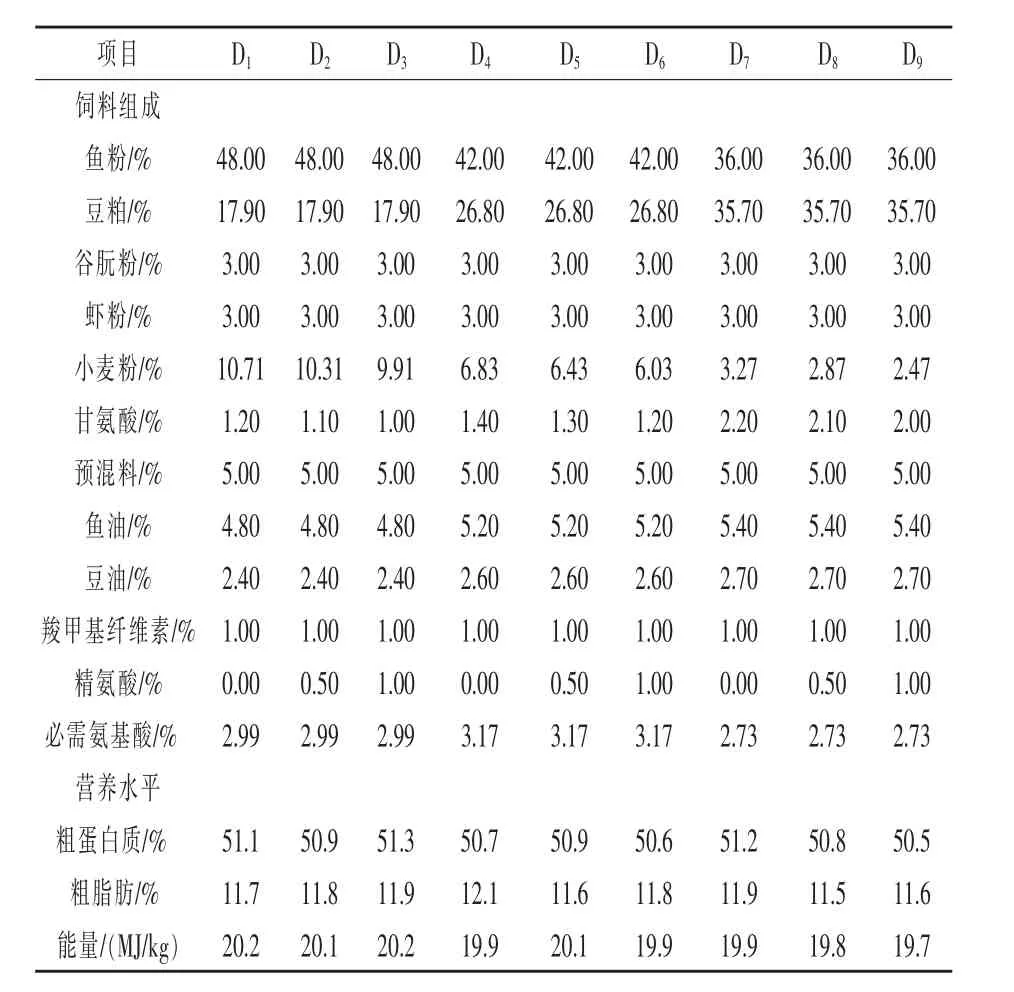
表1 试验饲料组成及营养水平

表2 鱼体及饲料原料氨基酸含量(干物质基础)%
1.3 样本采集 养殖试验结束时,试验鱼禁食24 h,每个箱中随机取5尾鱼,解剖取胃、前肠、中肠及后肠,每组随机选取前、中、后肠各3个样品用0.85%的生理盐水清洗后分别置于Bouin氏液中固定,8 h后取出移入70%乙醇中保存以用于后续肠道组织结构分析,剩余的样品分装于相应编号的密封袋,迅速转入-80℃低温冰箱内保存,以用于消化酶活力的测定。
1.4 试验方法
1.4.1 粗酶液的制备 将采样的各种组织在冰生理盐水中洗净,用滤纸吸干组织表面生理盐水,置于离心管内,用移液管移入9倍于组织块质量的0.85%生理盐水,匀浆,在4℃下4500 r/min离心15min,取上层清液备用。
1.4.2 生化指标检测 采用试剂盒 (南京建成生物工程研究所)对肠道、胃中的蛋白酶活性、淀粉酶活性和脂肪酶活性进行测定。
1.4.3 肠道结构的观察 将存放于70%乙醇中的组织取出,叔丁醇逐级脱水,常规石蜡包埋切片,厚度5~6μm,H·E染色,中性树胶封片。采用Motic BA600-4数字切片扫描与应用系统高精度扫描切片并保存。之后用Motic数字切片助理Lite 1.0观察组织并进行测量,每张切片测定10个视野,以得到的均值作为测定数据进行分析。
1.5 数据统计 所有数据均以“平均值±标准差”表示,并利用SPSS 18.0统计软件进行双因素方差分析(two-way ANOVA),当有交互作用时,用单因素方差分析(one-way ANOVA),若差异达到显著水平(P<0.05),则进行Tukey法多重比较。
2 结果
2.1 豆粕替代鱼粉并添加精氨酸对点带石斑鱼组织消化酶活性影响
2.1.1 豆粕替代鱼粉并添加精氨酸对点带石斑鱼蛋白酶活性的影响 由表3可以看出,豆粕替代水平和精氨酸添加水平的交互作用对点带石斑鱼前肠和后肠的蛋白酶活性影响显著(P<0.05),对点带石斑鱼中肠、胃的蛋白酶活性无明显影响(P>0.05);豆粕替代水平对后肠蛋白酶活性影响显著(P<0.05),对前肠、中肠、胃蛋白酶活性影响不显著(P>0.05);精氨酸添加水平对前肠、中肠蛋白酶活性影响显著(P<0.05),对后肠、胃蛋白酶活性影响不显著(P>0.05)。在精氨酸添加量为0%时,后肠蛋白酶活性随着替代水平的升高呈现先上升后下降的趋势,30%豆粕替代组比20%替代组提高了85.1%,比40%豆粕替代组提高了174%;40%替代组则比20%替代组降低了48%,在0.5%和1%精氨酸添加组中,随豆粕替代水平的升高,后肠蛋白酶活性在各组间无显著差异(P>0.05)。在20%豆粕替代组中,0.5%精氨酸添加组较0%和1%组分别提高了56.7%和54.4%(P<0.05),但是30%和40%豆粕替代水平间,各组别差异不显著(P>0.05)。
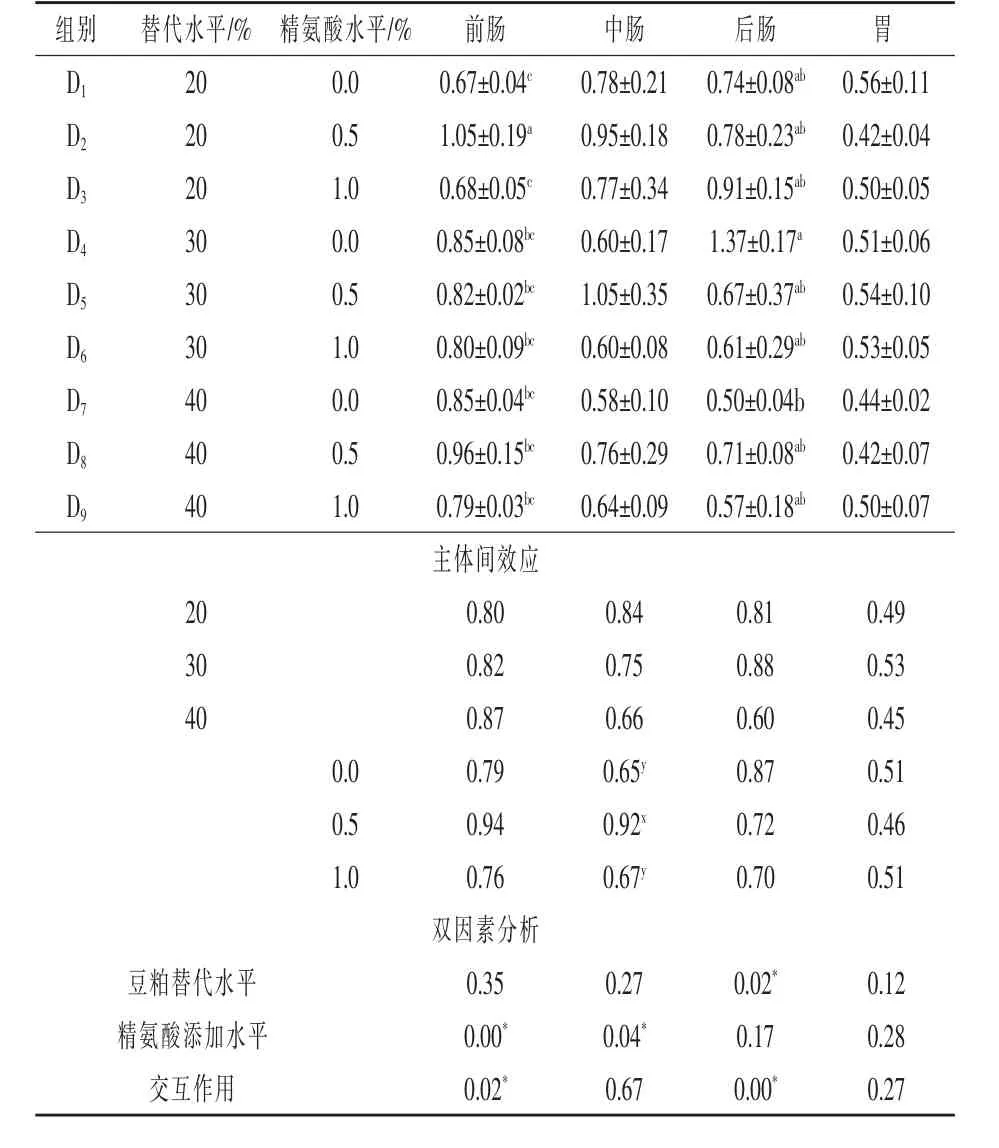
表3 豆粕替代鱼粉并添加精氨酸对点带石斑鱼蛋白酶活性的影响U/g prot
2.1.2 豆粕替代鱼粉并添加精氨酸对点带石斑鱼淀粉酶活性的影响 由表4可以看出,豆粕替代水平和精氨酸添加水平的交互作用显著影响了点带石斑鱼中肠淀粉酶活性(P<0.05),对前肠、后肠、胃淀粉酶无显著影响(P>0.05),在D9组(豆粕替代水平40%,精氨酸添加水平1%)时,中肠淀粉酶活性达到最高。豆粕替代水平和精氨酸补充水平对前肠、中肠、后肠的淀粉酶活性均无显著影响(P>0.05),精氨酸添加水平对胃淀粉酶活性无显著影响(P>0.05),而豆粕替代水平对胃淀粉酶活性有显著影响(P<0.05)。随豆粕替代水平增加,胃淀粉酶活性降低,同一精氨酸补充水平下,40%替代组胃淀粉酶活性显著低于20%替代组及30%替代组(P<0.05),20%和30%替代组胃淀粉酶活性没有显著差异(P>0.05),当精氨酸添加量为1%时,30%替代组胃淀粉酶活性比20%替代组降低了27.5%,40%替代组比20%替代组降低了72.5%。精氨酸添加水平对各组织淀粉酶活性无显著影响(P > 0.05)。

表4 豆粕替代鱼粉并添加精氨酸对点带石斑鱼淀粉酶活性的影响 U/gprot
2.1.3 豆粕替代鱼粉并添加精氨酸对点带石斑鱼脂肪酶活性的影响 由表5可知,豆粕替代水平和精氨酸添加水平的交互作用对后肠脂肪酶活性存在显著影响,对其他组织无明显影响 (P>0.05)。豆粕替代水平对后肠脂肪酶活性存在显著影响(P<0.05),对前肠、中肠、胃脂肪酶活性无显著影响(P>0.05)。随豆粕替代水平升高,后肠脂肪酶活性显著升高,在40%替代组最高,在0%、0.5%精氨酸添加水平下分别比30%替代组提高45.2%、81.3%(P>0.05),均显著高于20%替代组(P<0.05);而随着精氨酸添加水平的增加,后肠脂肪酶活性先上升后显著下降,当替代水平为40%,精氨酸添加水平为0.5%时,后肠脂肪酶活性达到最佳。精氨酸添加水平显著影响了中肠的脂肪酶活性(P<0.05),随精氨酸添加水平升高,中肠脂肪酶活性显著升高(P<0.05),当豆粕替代水平为40%,精氨酸添加水平为1%时,中肠的脂肪酶活性最佳;豆粕替代水平和精氨酸添加水平均对前肠和胃的脂肪酶活性没有显著影响(P>0.05)。

表5 豆粕替代鱼粉并添加精氨酸对点带石斑鱼脂肪酶活性的影响U/g prot
2.2 豆粕替代鱼粉并添加精氨酸对点带石斑鱼肠道结构的影响
2.2.1 豆粕替代鱼粉并添加精氨酸对点带石斑鱼前肠结构的影响 由表6可看出,豆粕替代水平和精氨酸添加水平以及两者的交互作用均对点带石斑鱼前肠的肌层厚度和皱襞高度存在显著影响(P<0.05),对点带石斑鱼前肠的黏膜下层厚度和上皮细胞宽度无明显影响(P>0.05)。随豆粕替代水平的升高,肌层厚度和皱襞高度均先升高后降低,在30%替代组最高,与20%替代组无明显差异(P > 0.05),均显著高于 40%替代组(P < 0.05)。 随精氨酸添加水平升高,肌层厚度显著升高 (P<0.05);而随精氨酸添加水平升高,肌层厚度和皱襞高度显著增加(P<0.05)。黏膜下层厚度和上皮细胞宽度不受替代水平和精氨酸添加水平的影响,黏膜下层厚度最大值发生在豆粕替代水平为20%,精氨酸添加水平为1%时,上皮细胞宽度在各处理组之间没有显著差异(P>0.05)。

表6 豆粕替代鱼粉并添加精氨酸对点带石斑鱼前肠结构的影响 μm
2.2.2 豆粕替代鱼粉并添加精氨酸对点带石斑鱼中肠结构的影响 由表7可看出,豆粕替代水平和精氨酸添加水平的交互作用对点带石斑鱼中肠的肌层厚度、上皮细胞宽度和皱襞高度存在显著影响(P<0.05),其中,豆粕替代水平显著影响了中肠肌层厚度(P<0.05),对其他指标无显著影响(P>0.05);而精氨酸添加水平对中肠肌层厚度和皱襞高度存在显著影响 (P<0.05),对黏膜下层厚度和上皮细胞宽度没有显著影响(P>0.05)。随豆粕替代水平的升高,肌层厚度先升高后降低,在30%替代组最高,与20%替代组无明显差异 (P>0.05),显著高于40%替代组 (P<0.05)。随精氨酸添加水平升高,肌层厚度和皱襞高度先升高后趋于平稳,0.5%和1%精氨酸添加组显著高于未添加组(P<0.05),在20%、40%豆粕替代水平组,精氨酸添加水平对中肠褶皱高度无显著影响(P>0.05),而在30%豆粕替代组,随精氨酸添加水平的升高皱襞高度先升高后降低,0.5%添加组比对照组提高15.8%(P<0.05),黏膜下层厚度和上皮细胞宽度在各处理组之间均没有显著差异(P>0.05)。

表7 豆粕替代鱼粉并添加精氨酸对点带石斑鱼中肠结构的影响 μm
2.2.3 豆粕替代鱼粉并添加精氨酸对点带石斑鱼后肠结构的影响 由表8可看出,豆粕替代水平和精氨酸添加水平的交互作用对点带石斑鱼后肠的黏膜下层厚度、肌层厚度、上皮细胞宽度和皱襞高度均不存在显著影响(P>0.05)。豆粕替代水平显著影响了后肠的肌层厚度和皱襞高度 (P<0.05),而精氨酸添加水平则显著影响了后肠黏膜下层厚度、肌层厚度和皱襞高度(P<0.05)。随豆粕替代水平的升高,肌层厚度降低,20%替代组高于40%替代组17.0%(P<0.05),与30%替代组无明显差异(P>0.05),在各处理组之间,肌层厚度最高值发生在替代水平为20%,精氨酸添加水平为1%组;而随豆粕替代水平的升高,皱襞高度先升高后下降,在30%替代组最高,与20%替代组无明显差异(P>0.05),高于40%替代组7.4%(P<0.05),皱襞高度在各处理组之间的最高值发生在30%豆粕替代水平,1%精氨酸添加水平组。随精氨酸添加水平升高,黏膜下层厚度、肌层厚度、皱襞高度均显著升高,最高值均发生在1%添加组,与0.5%添加组无明显差异(P>0.05),较0%添加组分别提高29.4%、16.4%、6.8%(P<0.05), 黏膜下层厚度最高值发生在替代水平为20%,精氨酸添加水平为1%的处理组。上皮细胞宽度在各处理组之间没有显著差异(P>0.05)。

表8 豆粕替代鱼粉并添加精氨酸对点带石斑鱼后肠结构的影响 μm
3 讨论
3.1 豆粕替代鱼粉并添加精氨酸对点带石斑鱼组织消化酶活性的影响 鱼类对营养物质消化和吸收的能力取决于鱼类消化酶活性,因此,对鱼类消化酶活性的研究有着非常重要的意义。在对大菱鲆幼鱼(李宗升,2016)和埃及胡子鲇(吴莉芳等,2010)的研究中发现随豆粕替代水平的提高,鱼体肠、胃蛋白酶活性呈下降趋势,对日本鲈鱼幼鱼(Li等,2014)的研究中,也发现蛋白酶活性随饲料中植物蛋白替代比例的上升而减小,认为导致蛋白酶活性下降的重要因素是植物蛋白中普遍含有的植酸。而在本试验中,豆粕替代量超过30%时,后肠蛋白酶及胃淀粉酶活性显著降低,试验结果与其相符,可能是豆粕作为植物蛋白源在替代鱼粉的过程中,当豆粕替代鱼粉比例过大,大豆中含有的一些抗营养因子,如皂甙、胰蛋白酶抑制因子和大豆凝集素等影响了鱼类对植物蛋白的吸收利用(邹文超等,2014)。此外,点带石斑鱼为肉食性,消化道中蛋白酶活性与其食性相适应,对植物蛋白的适应程度较低,且不及动物蛋白,因此,当豆粕替代水平过高时,蛋白酶活性下降,在黑鲷(杨彬彬等,2015)和翘嘴红鲌(钱曦等,2007)的相关研究中也有相似报道。同时,精氨酸添加水平为0.5%时,点带石斑鱼前肠、中肠的蛋白酶活性,以及鱼体后肠脂肪酶活性显著提高;在精氨酸添加水平为1%时,中肠肠脂肪酶活性显著提高。有研究发现,在黑鲷鱼幼鱼饲料中添加外源精氨酸能提高胃和前肠的蛋白酶和淀粉酶活力,从而提高黑鲷鱼饲料的消化率和吸收率(周凡,2011),以及在对杂交鲟鱼幼鱼 (Acipenser schrenckii×Acipenser baeri)消化酶活性的研究中,也发现适宜的精氨酸添加水平显著提高了幼鱼前肠的蛋白酶和淀粉酶活力、中肠蛋白酶活力 (王连生等,2017)。本试验结果与其相近,说明在饲料中添加适当水平的精氨酸可以提高点带石斑鱼肠道内消化酶的活性,有利于肠道对营养物质的消化吸收。
3.2 豆粕替代鱼粉并添加精氨酸对点带石斑鱼肠道结构的影响 肠道是鱼类营养物质消化和吸收的器官,主要由黏膜层、黏膜下层、肌肉层和浆膜层四层结构构成,其中由黏膜层和黏膜下层向肠腔突起形成的肠道皱襞,其高度可反映该鱼体肠道吸收面积,如对大西洋白姑鱼 (Ribeiro等,2014)、美国红鱼(Zhou 等,2010)、鲤(Geda 等,2012)等的研究中将皱襞的高低作为动物肠道吸收能力的重要指标。有研究表明,在大西洋鲑(Krogdahl等,2015)、 欧洲真鲈 (Kaushik 等,2004)、斑马鱼幼鱼(Hedrera 等,2013)和黄金鲈(吴莉芳等,2017)的饲料中添加一定量的大豆产品后,会导致鱼后肠组织结构发生形态变化,对后肠产生不利影响,使其组织结构完整性遭到破坏,如上皮细胞与固有层分离、肌层变薄、肠绒毛脱落或破损现象。在本试验中,随豆粕替代水平的升高,后肠的肌层厚度降低,在20%替代组最高,且皱襞厚度先升高后下降,在30%替代组最高,此试验结果与以上研究结果相符,说明高豆粕替代水平在一定程度上对鱼体肠道造成了不利影响。从而影响了鱼体对营养物质的消化吸收,降低了鱼体的生长,本试验中补充了豆粕替代鱼粉后引起的必需氨基酸不平衡,但是氨基酸的消化吸收率没有测定,也许氨基酸的吸收不同步以及豆粕中的抗营养因子如大豆凝集素、大豆抗原蛋白都可能引起鱼类肠道对营养物质的吸收以及皱襞高度的下降。
在对美国红鱼的研究中发现,1%精氨酸可提高鱼体前肠和中肠皱襞高度以及前肠的上皮细胞高度 (Cheng等,2011),对杂交条纹鲈(Morone chrysops×Morone saxatilis)的研究表明,添加1%~2%外源精氨酸能提高肠道绒毛高度、肠上皮细胞高度和微绒毛高度(Cheng等,2012)。同样,在饲料中添加适宜水平的精氨酸能显著提高杂交鲟幼鱼前肠和中肠的皱襞高度、前肠肌层厚度(王连生等,2017),对黄颡鱼的研究显示,饲料中精氨酸水平的增加可以有效提高肠道肌层厚度和皱襞高度。在本试验中,随精氨酸添加水平升高,前中后肠的肌层厚度、皱襞高度均升高,这与在美国红鱼和杂交条纹鲈上的研究结果一致 (Cheng等,2012、2011),而在对斜带石斑鱼的研究中发现,随精氨酸添加水平的升高前肠后肠的肌层厚度、皱襞高度以及中肠的肌层厚度均呈现先增高后下降的趋势,且在2.95%添加组达到最高 (迟淑艳等,2016),与本试验结果略有不同,本试验中随着精氨酸添加水平的升高鱼体肠道的肌层厚度、皱襞高度均升高,可能是本试验精氨酸的添加量还不足以使肠道肌层厚度和皱襞高度等出现抑制现象。这些都说明在饲料中适当替代豆粕和添加适当水平的精氨酸有利于肠道对营养物质的消化吸收。
4 结论
在本试验条件下,当豆粕蛋白替代鱼粉蛋白不超过30%,精氨酸添加量为0.5%~1%,对石斑鱼的消化和肠道结构有利。
猜你喜欢
体育科技文献通报(2022年4期)2022-10-21
当代水产(2022年8期)2022-09-20
当代水产(2022年6期)2022-06-29
当代水产(2022年1期)2022-04-26
中国动物保健(2021年8期)2021-11-24
当代水产(2020年3期)2020-06-15
当代水产(2019年7期)2019-09-03
当代水产(2019年6期)2019-07-25
当代水产(2019年5期)2019-07-25
当代水产(2019年1期)2019-05-16
