
胰腺癌组织中PSAT1表达及其介导的细胞增殖 侵袭作用机制研究*
2019-01-23 06:48聂钊李岚杨兰群崔德军钱利叶丽敏杨倩张德林褚明亮曾宪春
中国肿瘤临床 2018年23期
聂钊 李岚 杨兰群 崔德军 钱利 叶丽敏 杨倩 张德林 褚明亮 曾宪春
胰腺癌是一种恶性程度较高的消化系统肿瘤,根据中国国家癌症中心发布的最新数据,我国胰腺癌发病率居恶性肿瘤第10位,死亡率居第5位[1]。尽管近20年癌症诊疗技术得到了快速发展,但是胰腺位置较深,加之起病隐匿,早期诊断胰腺癌仍然是临床的难题之一。当出现临床症状时往往已是中晚期,患者5年生存率低于8%[2-3]。因此研究胰腺癌的发病机制,寻找可靠的早期诊断和预后评估标志物,是改善胰腺癌预后的关键。高糖酵解水平(Warburg效应)是恶性肿瘤的重要特征[4],肿瘤细胞通过Warburg效应为细胞生长和增殖提供足够的能量。研究证实[5],Warburg效应能诱导一系列编码糖酵解代谢酶的基因表达,进而促进细胞生物量累积和肿瘤细胞增殖。磷酸丝氨酸转氨酶1(phosphoserine aminotransferase 1,PSAT1)是丝氨酸合成的限速酶,在3-磷酸甘油酸酯转化为丝氨酸过程中发挥关键作用[6]。在这一转化过程中,丝氨酸能为肿瘤细胞异常生长和增殖提供充足的能量供应,因此PSAT1与肿瘤细胞的发生和进展密切相关。PI3K/Akt/mTOR通路是细胞物质、能量代谢主要信号通路,在肿瘤细胞生长和增殖中发挥重要作用,PI3K/Akt/mTOR通路也被认为是肿瘤发生的主调节器[7-8]。本研究通过检测胰腺癌组织和细胞PSAT1表达水平,并从PI3K/Akt/mTOR通路角度出发,初步探讨PSAT1在胰腺癌增殖、侵袭中的作用及机制。
1 材料与方法
1.1 材料
1.1.1 组织样本 采集2013年7月至2017年7月在贵州省人民医院行手术切除的98例胰腺癌患者组织样本,包括癌组织和配对的癌旁组织(距肿瘤切缘≥1 cm);样本组织取出后立即置于液氮中冷冻,-80℃保存供后续实验使用。所有患者术前完善双源CT胰腺增强扫描,并经病理学证实为胰腺腺癌。术前未进行化疗、放疗或其他抗癌治疗,排除转移性胰腺癌、伴有其他恶性肿瘤或自身免疫性疾病者。本研究经医院伦理委员会批准(编号:GZRY20130513),所有患者术前均签署知情同意书。
1.1.2 细胞来源与培养 人胰腺癌细胞系BxPC-3、PANC1、SW1990细胞购自美国典型培养物保藏中心(ATCC),正常胰腺腺泡细胞AR42J购自中国科学院上海细胞库。BxPC-3、SW1990、PANC1细胞接种于DMEM培养基,AR42J细胞接种于RPMI-1640。DMEM培养基和RPMI-1640培养基均含有10%胎牛血清(FBS)、100 U/mL青霉素和链霉素,在细胞培养箱中以37℃、5%CO2条件培养。
1.1.3 实验试剂 DMEM培养液、胎牛血清(美国Gibco公司),RPMI-1640培养液、Lipofectamine 2000脂质体、Opti-MEM、TRIzol试剂(美国Invitrogen公司),PSAT1-siRNA(上海吉玛制药技术有限公司),Cell Counting Kit-8(Dojindo公司),Transwell小室(康宁公司),RIPA裂解液、BCA试剂(碧云天生物技术研究所),ECL检测试剂盒(Thermo scientific公司),鼠抗人PSAT1单克隆抗体、鼠抗人Akt、mTOR、p-Akt、pmTOR单克隆抗体(美国Abcam公司),辣根过氧化物酶标记山羊抗鼠IgG(武汉博士德生物工程公司)。
1.2 方法
1.2.1 免疫组织化学染色检测组织PSAT1表达 石蜡包埋的样本组织经二甲苯脱蜡,梯度乙醇水化;加入柠檬酸盐缓冲液(pH=6.0)加热30 min修复抗原,3%H2O2孵育10 min消除内源性过氧化物酶,加入5%山羊血清封闭2 h。样本组织加入PSAT1抗体4℃孵育过夜,次日再加入二抗孵育1 h,光学显微镜下观察染色结果。结果判定:按照染色强度和阳性细胞染色面积对PSAT1表达进行综合评价,染色强度:0分,阴性;1分,弱染色;2分,中等染色;3分,强染色。染色面积:0分,0~5%;1分,6%~25%;2分,26%~50%;3分,>50%;最终评分为染色强度评分×染色面积。参考 De Marchi等[9]报道,最终评分≥3 分为PSAT1表达阳性,<3分为PSAT1表达阴性。
1.2.2 临床资料整理与术后随访 收集患者临床资料,包括性别、年龄、手术时肿瘤大小、分化程度、肿瘤位置、淋巴结转移、TNM分期、神经侵犯情况等。患者术后进行规律随访,本研究随访截止日期为2018年4月30日,随访时间5~53个月,中位随访时间17.3个月。记录患者总生存期和无病生存期,定义总生存期为术后第2 d至死亡或末次随访的时间,无病生存期为术后第2 d至胰腺癌首次复发或转移的时间。
1.2.3 小干扰RNA(siRNA)瞬时转染 利用siRNA技术敲低细胞PSAT1表达,同时以转移无义序列的细胞作为阴性对照(NC-siRNA)。转染前24 h将细胞以1×105个/孔接种于6孔板,待细胞50%~70%融合时,根据Lipofectamine 2000说明书,siRNA和Lipofectamine 2000分别加入250 μL Opti-MEM培养液稀释,室温静置5 min后轻轻混匀,再将500 μL混合物滴加至6孔板,转染48 h后进行后续实验。PSAT1-siRNA序列:正义5′-CGC CAAGAAGTTTGGGACTATAT-3′,反义5′-TATAGTCCC AAACTTCTTGGCT-3′。
1.2.4 CCK-8检测细胞增殖活性 取对数生长期细胞,以2×103个/孔接种于96孔板,37℃、5%CO2条件培养箱中培养。分别于细胞培养24、48、72、96 h时向每孔中加入10 μL CCK-8试剂,继续培养1.5 h,酶标仪读取波长为450 nm的吸光度值,并通过吸光度值绘制细胞生长曲线。每组平行设置5个复孔,实验重复3次,取平均值。
1.2.5 细胞划痕实验 取对数生长期细胞,以5×105个/孔接种于6孔板,加入不含血清的DMEM培养基培养至100%融合。用10 μL移液枪头在细胞层划“一”字划痕,PBS冲洗3遍,再加入含胎牛血清的DMEM培养基继续培养24 h,显微镜下观察细胞迁移的距离,并计算细胞迁移率。
1.2.6 Transwell侵袭实验 细胞用不含血清培养基稀释,计数,配成细胞悬液;将1×105个细胞溶于100 μL DMEM培养液,置于Transwell上室,下室中加入800 μL含有10%FBS的条件培养基。37℃孵育24 h,取出上室,用PBS洗2遍,5%戊二醛4℃固定。加入0.1%结晶紫染色,室温染色10 min,PBS洗2次,用棉球擦去上表面细胞,显微镜下取5个随机视野计穿过滤膜的细胞数,统计结果。
1.2.7 Western blot待细胞生长至80%~90%融合时,加入RIPA裂解液充分裂解细胞,提取总蛋白,并利用BCA试剂盒进行蛋白定量。30 μg总蛋白用SDS-PAGE凝胶电泳分离,电转至PVDF膜,然后加入含有5%脱脂奶粉的TBST液室温封闭1.5 h,PBS冲洗 2 遍,加入 PSAT1(1:200)、Akt(1:500)、mTOR(1:300)、p-Akt(1:1 000)、p-mTOR(1:1 000)、GAPDH(1:1 000)一抗,4℃孵育过夜。次日PBS冲洗3遍,加入辣根过氧化物酶(HRP)标记山羊抗鼠二抗(1:1 000)室温孵育1h,PBS洗涤后ECL显影,以GAPDH为内参,Image J扫描蛋白灰度值。目的蛋白相对灰度值=目的蛋白灰度值/GAPDH灰度值,结果以实验组相对灰度值/正常对照组相对灰度值表示实验组蛋白相对表达量。
1.3 统计学方法
使用SPSS 19.0软件、GraphPad Prism 7.0软件和和Adobe Photoshop CS 6软件进行数据和图形处理。计数资料用百分比表示,χ2检验分析PSAT1表达与胰腺癌患者临床资料的关系。计量资料用x±s表示,多组间比较采用单因素方差分析(One-way ANOVA),两组间比较采用Student′st检验。Kaplan-Meier法绘制胰腺癌患者生存曲线,各组间生存率比较采用Log-rank法;多因素采用Cox比例风险模型。以P<0.05表示差异具有统计学意义。
2 结果
2.1 胰腺癌组织和癌旁组织PSAT1表达
PSAT1表达主要定位于细胞质或细胞浆,PSAT1在胰腺癌组织中阳性表达率为69.4%(68/98),明显高于癌旁组织5.0%(5/98),两者比较差异具有统计学意义(χ2=86.638,P<0.001,图1)。
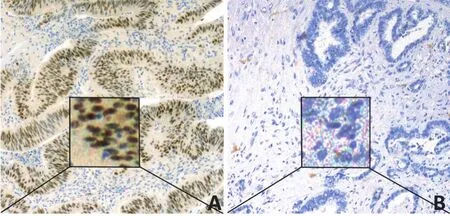
图1 胰腺癌组织和癌旁组织中PSAT1表达
2.2 PSAT1表达与胰腺癌临床资料的关系
98例胰腺癌患者中,PSAT1表达阳性率与淋巴结转移、TNM分期有关(P<0.05),与年龄、性别、分化程度、肿瘤位置、神经侵犯等无关(P>0.05,表1)。
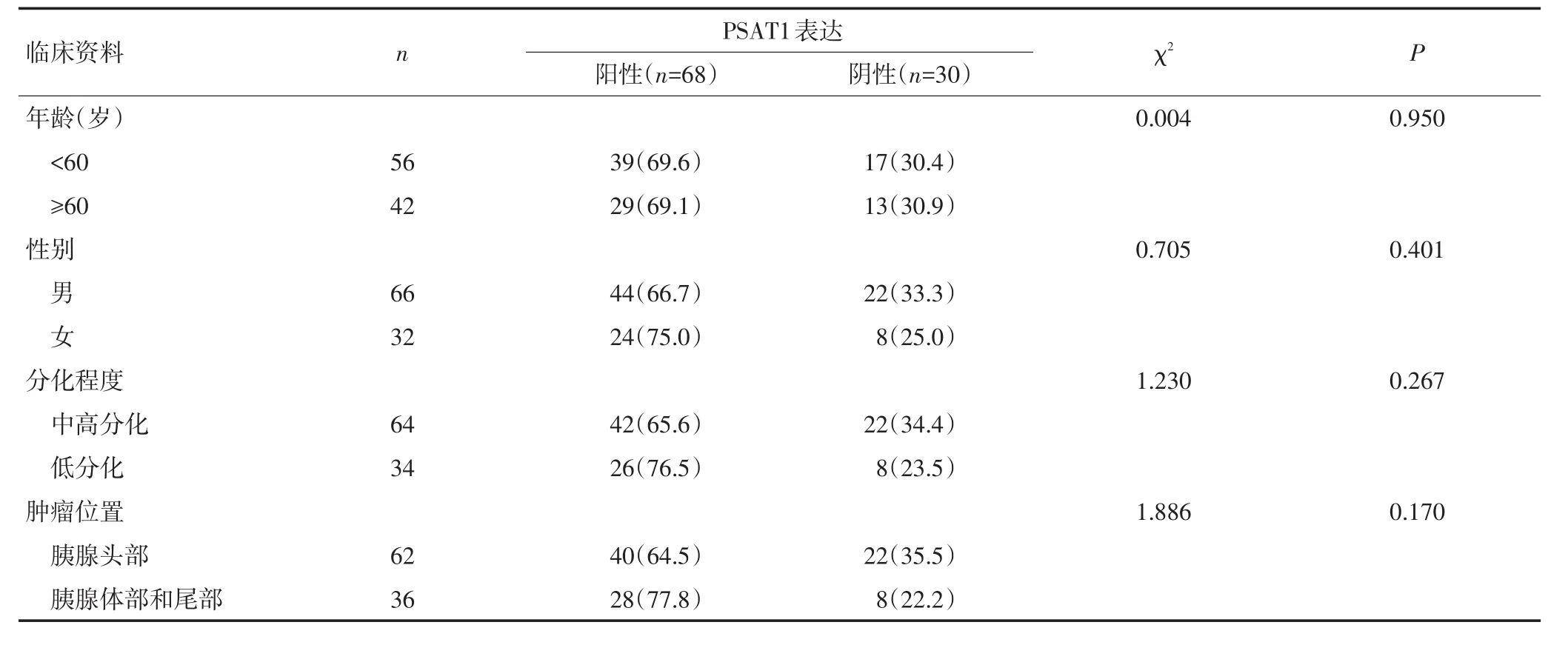
表1 PSAT1表达与胰腺癌临床资料的关系 n(%)

表1 PSAT1表达与胰腺癌临床资料的关系 n(%)(续表1)
2.3 PSAT1表达与胰腺癌预后的关系
生存分析结果显示,PSAT1高表达的胰腺癌患者总生存期和无病生存期明显低于PSAT1低表达患者(P<0.05,图2)。Cox多因素回归显示,PSAT1表达是影响胰腺癌总生存的危险因素(P<0.05),PSAT1表达同时也是影响胰腺癌无病生存的危险因素(P<0.05,表2)。
2.4 抑制PSAT1表达对胰腺癌细胞增殖的影响
由图3A可见,胰腺癌细胞系BxPC-3、PANC1、SW1990中PSAT1蛋白表达明显高于AR42J细胞(P<0.05);其中PSAT1蛋白在BxPC-3、SW1990细胞中表达最高(2.17±0.18和2.28±0.20),因此选择BxPC-3、SW1990细胞进行后续功能学实验。向BxPC-3、SW1990细胞转染PSAT1-siRNA后,PSAT1-siRNA组PSAT1蛋白表达显著低于NC-siRNA组(P<0.05,图3B,3C),说明siRNA可以显著降低BxPC-3、SW1990细胞PSAT1表达。CCK-8实验结果显示,随着实验的延长,PSAT1-siRNA组细胞增殖速度减慢,在第72 h和96 h时,PSAT1-siRNA组细胞增殖率明显低于NC-siRNA组(P<0.05,图3D,3E),
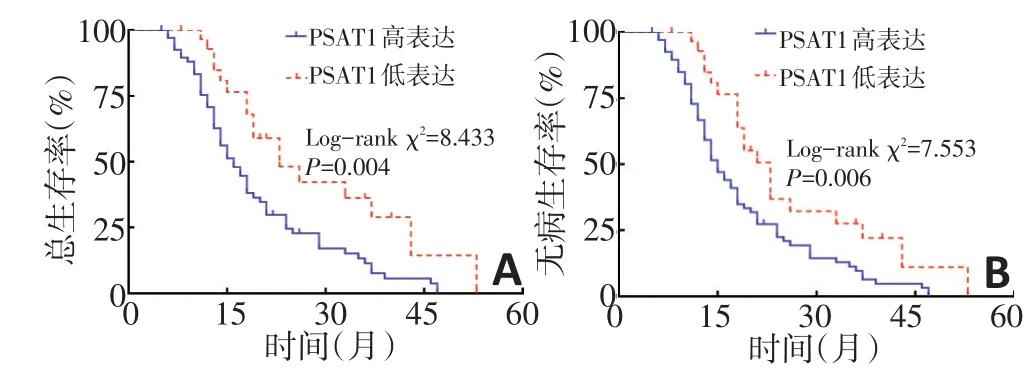
图2 不同PSAT1表达胰腺癌患者总生存曲线和无病生存曲线
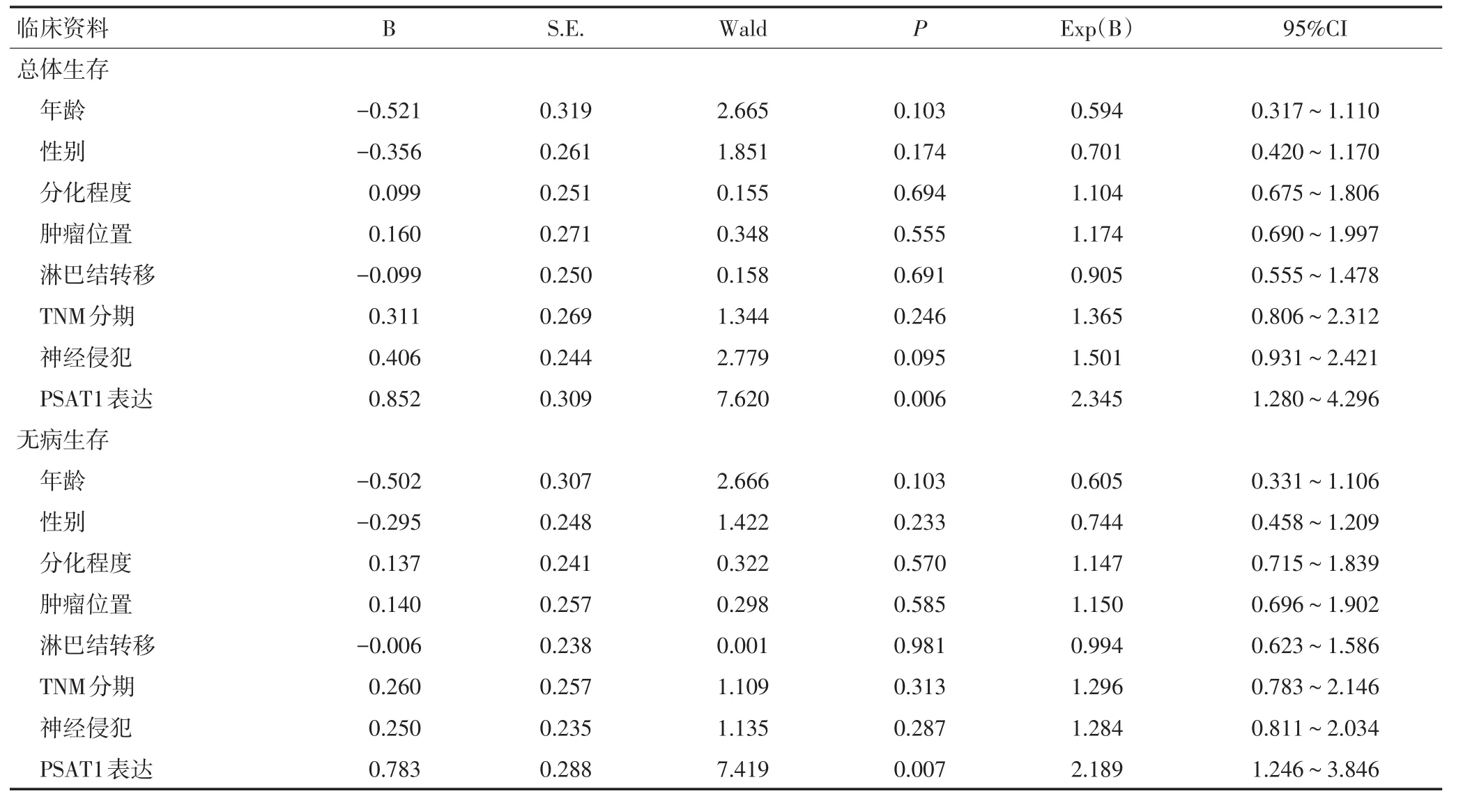
表2 Cox多因素分析影响胰腺癌总生存期和无病生存期的独立危险因素
2.5 抑制PSAT1表达对胰腺癌细胞迁移和侵袭的影响
细胞划痕实验结果显示,与NC-siRNA组比较,转染PSAT1-siRNA后BxPC-3、SW1990细胞相对迁移距离显著缩短,差异具有统计学意义(P<0.05,图4A,4B)。Transwell侵袭实验结果显示,PSAT1-siRNA组侵袭细胞数明显低于NC-siRNA组,差异具有统计学意义(P<0.05,图4C,4D)。
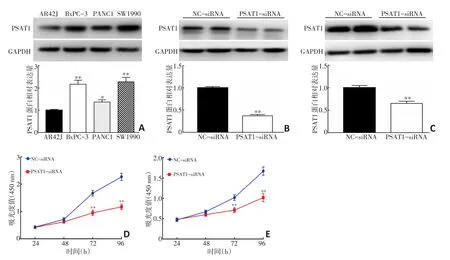
图3 抑制PSAT1表达对胰腺癌细胞增殖能力的影响
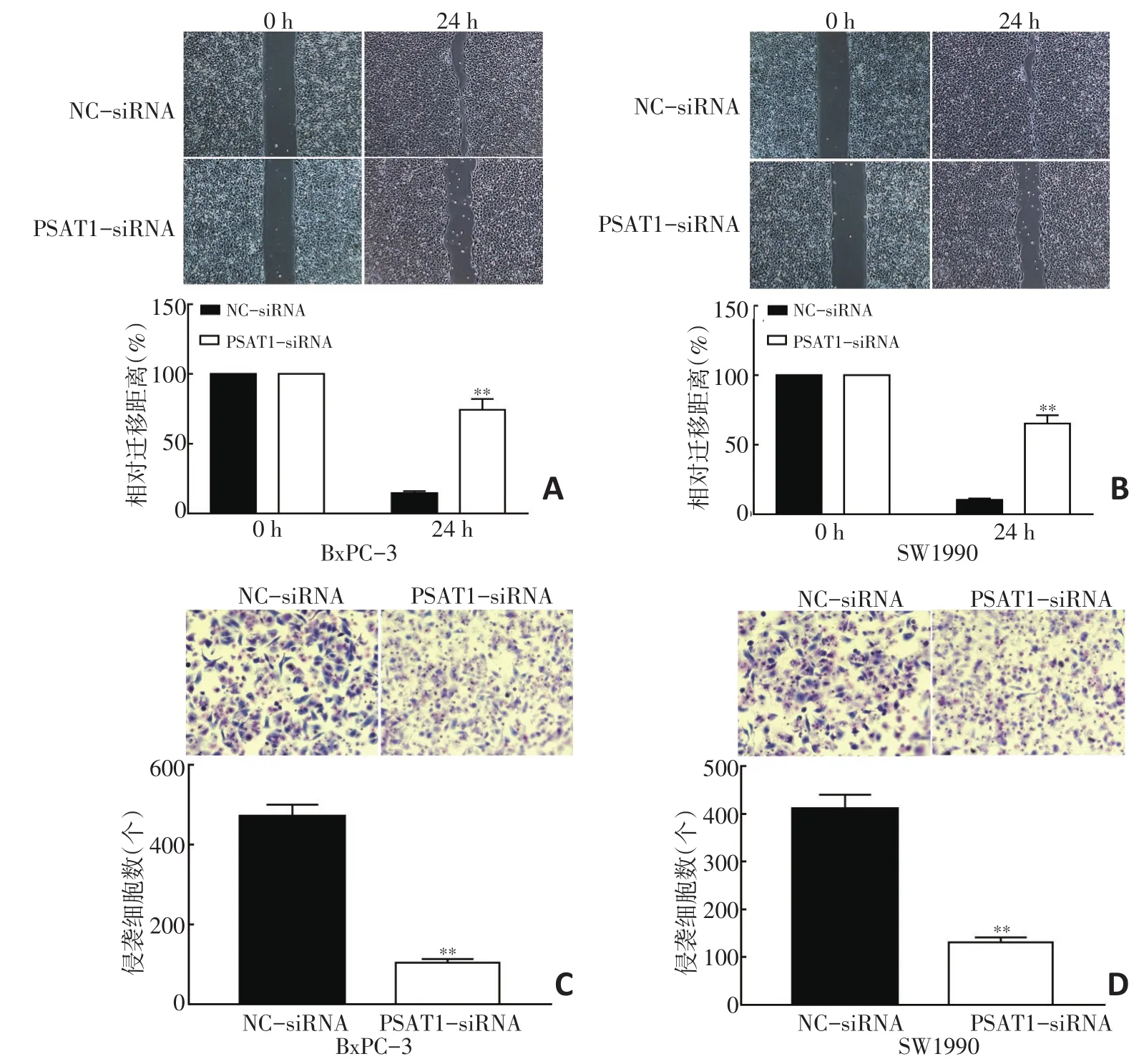
图4 抑制PSAT1表达降低胰腺癌细胞迁移和侵袭能力
2.6 抑制PSAT1表达对胰腺癌细胞PI3K/Akt/mTOR通路相关蛋白表达的影响
本研究利用Western blot检测BxPC-3和SW1990细胞PI3K/Akt/mTOR通路重要蛋白:Akt、mTOR蛋白及其活化形式p-Akt、p-mTOR的变化,结果显示,与NC-siRNA组相比,PSAT1-siRNA组p-Akt、p-mTOR蛋白表达显著降低(P<0.05,图5)。
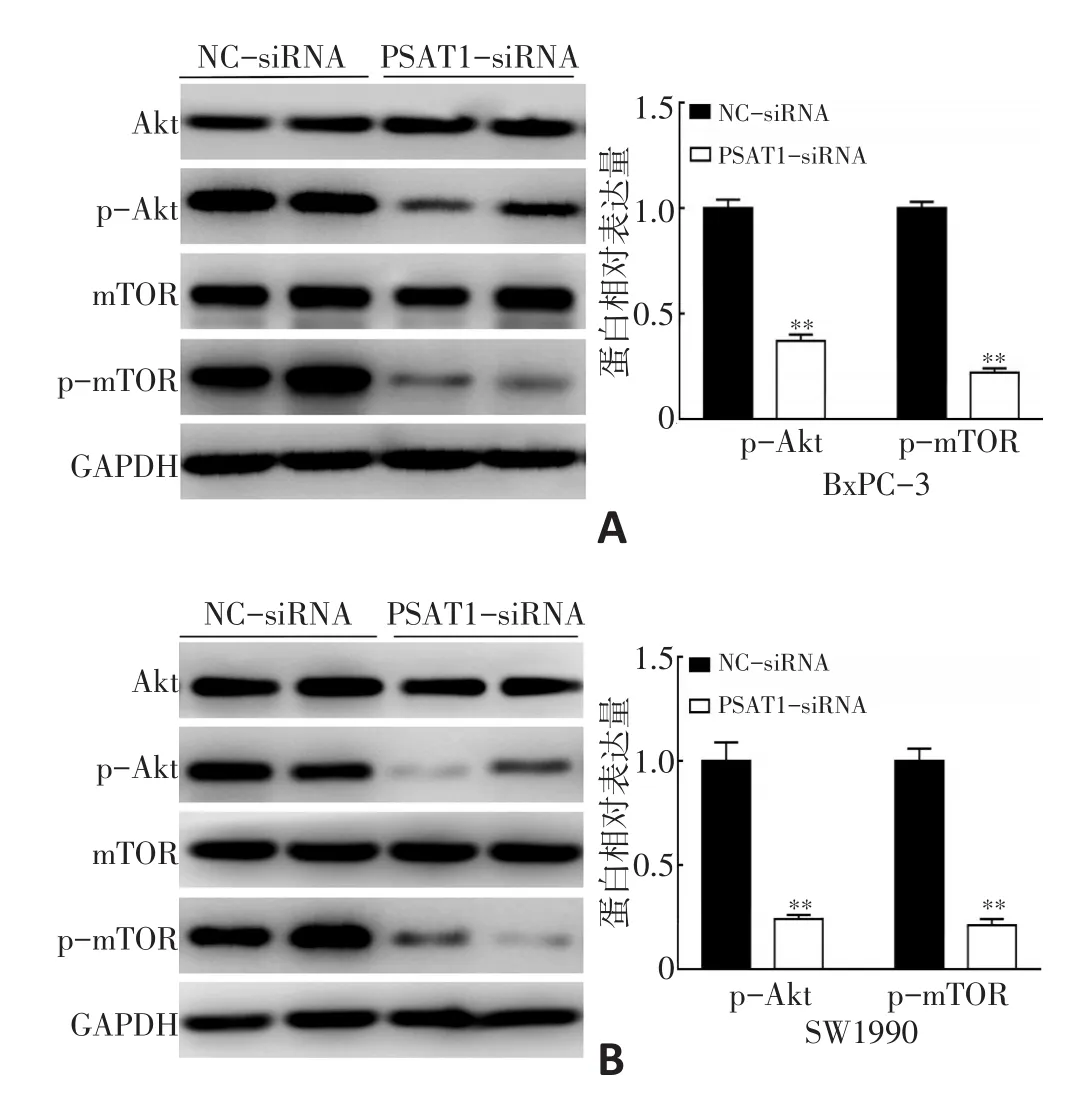
图5 抑制PSAT1表达对胰腺癌细胞Akt、mTOR蛋白表达的影响
3 讨论
代谢变化是肿瘤细胞典型特征之一,低效能的糖酵解途径(Warburg效应)为肿瘤细胞提供了大量的碳源,从而满足肿瘤快速生长的需要[10]。糖酵解途径还可以避免产生自由基,使肿瘤细胞逃避凋亡。因此,在糖酵解过程中发挥关键作用的代谢酶有望成为肿瘤诊疗的潜在靶点。PSAT1是磷酸丙酮酸氨基酸转移酶编码基因,在糖酵解中发挥重要作用。PSAT1是催化磷酸丝氨酸合成的限速酶,通过糖酵解3-磷酸甘油酸酯生成3-磷酸丙酮酸,将糖酵解产物引入丝氨酸合成途经,并为细胞提供原料和能量[11]。Qian等[12]报道PSAT1在结直肠癌组织中高表达,并且与肿瘤细胞对伊立替康、5-氟尿嘧啶和亚叶酸钙的敏感性有关。Gao等[13]也证实PSAT1通过调节ATF4促进乳腺癌细胞增殖,且高表达PSAT1与雌激素受体阴性乳腺癌不良预后有关。
Song等[14]证实糖酵解途径中另外一个关键酶磷酸甘油酸脱氢酶(PHGDH),与胰腺癌异常增殖和不良预后有关。基于此,推测PSAT1可能也参与了胰腺癌的多种生物学行为。为了验证此假设,本研究首先利用免疫组织化学染色检测胰腺癌组织和癌旁组织PSAT1表达,结果显示PSAT1在胰腺癌组织中阳性表达率明显高于癌旁组织,同时胰腺癌细胞PSAT1表达也高于胰腺细胞,说明胰腺癌中PSAT1异常表达。进一步分析发现,高表达PSAT1与胰腺癌肿瘤体积、淋巴结转移和TNM分期有关,提示PSAT1有作为早期诊断胰腺癌的价值。Liu等[15]报道PSAT1表达升高常提示发生食管鳞状细胞癌风险升高,并且PSAT1持续高表达可以作为患者不良预后的预测指标。Liao等[16]研究发现PSAT1表达与鼻咽癌瘤体体积、TNM分期、分化程度有关,与PSAT1低表达者比较,PSAT1高表达鼻咽癌患者局部无复发生存率、无远处转移生存率、疾病特异生存率和总体生存率均显著缩短。本研究对胰腺癌患者进行随访,结果发现SAT1高表达的胰腺癌患者总生存期和无病生存期明显低于PSAT1低表达患者,PSAT1表达是胰腺癌总体生存和无病生存的危险因素。上述结果说明胰腺癌组织PSAT1表达与胰腺癌发生和不良预后有关,PSAT1有望成为胰腺癌诊断和预后的评估指标。
Singh等[17]研究发现,复发性乳腺癌中PSAT1的高表达与细胞内丝氨酸水平呈正相关,沉默PSAT1表达可以降低丝氨酸水平,进而抑制细胞的生长和增殖。本研究利用siRNA技术沉默胰腺癌BxPC-3、SW1990细胞PSAT1表达,结果发现PSAT1基因沉默后,胰腺癌细胞增殖速度明显减慢,迁移和侵袭能力明显减弱,说明PSAT1与胰腺癌生长和远处转移有关。Hwang等[18]报道肿瘤细胞依靠丝氨酸生物合成途经为细胞快速增殖提供原料,抑制PSAT1可以阻断丝氨酸合成,导致细胞能量和原料供应不足,细胞分裂周期阻滞,最终引起细胞凋亡。Yang等[19]证实PSAT1基因沉默可以使非小细胞肺癌细胞周期调节蛋白cyclin D1降解,细胞分裂被阻滞于G0/G1期,最终抑制细胞增殖。
目前对PSAT1的研究尚处于起始阶段,其对肿瘤生物学行为的调控作用及相关机制仍未阐明。PI3K/Akt/mTOR通路是一条经典信号通路,在调节糖原代谢、细胞增殖中发挥重要作用[20]。研究显示[21],激活PI3K/Akt/mTOR通路能显著提高细胞的糖酵解水平,而PI3K抑制剂LY294002呈浓度、时间依赖性的抑制胃癌细胞增殖。在PI3K/Akt/mTOR通路中,活化的PI3K能激活下游关键蛋白Akt,使Akt作用其底物mTOR作用,完成调控细胞代谢、增殖和凋亡等作用[22]。Sharma等[23]报道胰腺癌细胞p-PI3K、p-Akt和p-mTOR蛋白表达明显上调,Mitofusin2可以通过抑制PI3K/Akt/mTOR信号通路诱导胰腺癌细胞自噬和线粒体融合。本研究对转染PSAT1-siRNA的BxPC-3、SW1990细胞PI3K/Akt/mTOR通路相关蛋白表达进行分析,结果显示与NC-siRNA组相比,PSAT1-siRNA组p-Akt、p-mTOR蛋白表达显著降低,提示沉默PSAT1可以抑制PI3K/Akt/mTOR通路的活性,PSAT1可能通过调节PI3K/Akt/mTOR通路参与胰腺癌细胞的增殖和侵袭。
综上所述,PSAT1在胰腺癌组织和细胞中呈高表达,PSAT1可能通过调节PI3K/Akt/mTOR通路参与胰腺癌细胞的增殖和侵袭。胰腺癌发病机制复杂,本研究仅单纯观察了PSAT1的变化,是否同时存在其他代谢基因改变尚不清楚。另外本组研究的样本量偏少,特别是分层之后更少,使得在临床观察中存在偏倚结果的可能。接下来的工作中,我们将进一步研究PSAT1对胰腺癌诊疗的意义以及相关的调控机制。
猜你喜欢
中国农业科学(2022年12期)2022-06-28
中国畜牧杂志(2022年1期)2022-01-20
华夏医学(2021年1期)2021-12-05
南昌大学学报(医学版)(2021年1期)2021-11-29
世界最新医学信息文摘(2021年25期)2021-05-13
食品与生物技术学报(2021年2期)2021-01-16
中国循证心血管医学杂志(2020年11期)2020-01-08
中成药(2019年12期)2020-01-04
中国现代医生(2016年31期)2017-03-02
中国现代医生(2016年32期)2017-03-02
