
耕作方式和施氮量对旱地冬小麦开花后干物质转运特征、糖含量及产量的影响
2019-01-21 02:58张礼军鲁清林张文涛白玉龙汪恒兴杨长刚
麦类作物学报 2018年12期
张礼军,鲁清林,张文涛,白玉龙,周 刚,汪恒兴,杨长刚
(1.甘肃省农业科学院小麦研究所,甘肃兰州 730070;2. 甘肃省天水市农业学校,甘肃清水 741400)
甘肃旱地冬小麦种植面积占全省小麦种植面积的60%以上,其多种植于西北黄土高原半干旱雨养区。该区域降水少,农业生产十年九旱,且常伴随较严重的土壤贫瘠[1-2],以致小麦产量长期低而不稳。传统翻耕模式对土壤结构破坏严重,加剧了水土流失,耗能过多,效益降低[3]。实施以覆盖(地膜和秸秆)为核心的免耕、少耕等保护性耕作技术能明显改善土壤理化性质,影响土壤的蓄水控温保肥能力,创造适合作物生长的外部环境,进而改变其内部同化过程,提高作物产量,有利于农业可持续性发展[4-7]。近年来,以全膜覆土穴播为代表的多茬免耕覆膜种植技术因有效地改善了土壤水、肥、气、热等环境,大幅度提高作物产量,在甘肃省旱地冬麦区得到大面积推广[2,8-9]。但不足的是持续的地膜种植造成“白色”污染[1-2],背离当前粮食绿色增产的要求。同时,由于种植小麦比较效益较低,农户很少将作物秸秆残茬等归还农田用于培肥地力[1]。高产的获得依赖过量施用氮肥,导致肥料利用率降低,地力退化[7]。因此,如何合理施用氮肥及控制地膜的使用,减少农业面源污染,实现西北黄土高原半干旱区作物的可持续生产,是该区域当前农业发展的重中之重。
研究表明,合理施用氮肥和采用保护性耕作技术均能大幅度提高旱地冬小麦籽粒产量,而二者有机结合的增产效果更显著,且有利于农业可持续发展[1,3,6]。小麦产量的高低既取决于开花后的光合生产能力,又决定于开花前碳水化合物的积累和开花后向籽粒的转运[10-11]。碳水化合物按其存在形式可分为结构性和非结构性碳水化合物。非结构性碳水化合物是植物生命活动的主要反应物质,以可溶性糖为主要形式暂贮于植株器官中。小麦籽粒产量与植株可溶性糖含量呈正相关,尤其与茎秆可溶性糖含量的相关性更高,但结果因环境变化而不同[12-15]。在非胁迫条件下,小麦营养器官中暂时贮存的碳水化合物对籽粒产量的贡献率为10%~20%[14],而在胁迫条件(高温、后期干旱、寡日照等)下可达50%~75%[14,16]。现有的研究主要集中在品种间可溶性糖的代谢差异及其对逆境胁迫(水分、氮素等)的响应方面[10-11,17-18],少有研究涉及到耕作方式和施氮量组合对旱地冬小麦灌浆期物质分配、糖代谢特征及其与产量关系的影响。基于此,本研究通过大田定位试验,分析了不同耕作方式和施氮量组合下旱地小麦花后干物质转运特征、糖含量及产量的差异,以期为西北黄土高原半干旱区冬小麦的高产、可持续栽培中合理耕作和施氮提供指导依据。
1 材料与方法
1.1 试验地概况
试验于2012-2015年在甘肃省农业科学院小麦研究所清水试验站进行。试验站海拔1 450 m,年降雨量500 mm左右,年均温9 ℃,属典型的西北黄土高原半干旱雨养农业区。试区土壤类型为黄沙土,耕层土壤的有机质含量1.121%,可溶性氮含量0.094 g·kg-1,有效磷含量16.39 mg·kg-1,速效钾含量97.22 mg·kg-1。
1.2 试验设计
试验于2012年开始定位实施,2014-2015年为该试验的第3年重复试验。试验采用双因素裂区设计。主区为耕作方式,分别为常规耕作(西北旱地冬小麦种植的常规模式,CT)、秸秆还田(翻耕秸秆粉碎还田,CTI)、全膜覆土穴播(PM)和免耕秸秆覆盖(NTS)。CT(对照模式):小麦收获时留茬10~15 cm,其余秸秆随籽粒带走,7月中旬用铧式犁深耕20 cm,9月下旬耙耱后播种,平作,条播,行距20 cm。CTI:前茬作物收获后,结合传统耕作将长度约5 cm的秸秆4 500 kg·hm-2翻入土壤,平作,条播,行距20 cm,以后每季将收获秸秆全部还田。PM:在前茬小麦收获后,秸秆带走并进行翻耕,种植前覆盖地膜,并在膜上覆盖1 cm厚度的干土,穴播,行距20 cm,穴距12 cm;该模式因其增产潜力高,并且克服了其他地膜覆盖的缺点,近年来在甘肃省旱地冬麦区得到大面积推广。NTS:前作收获后,地表均匀覆盖长度5 cm的秸秆4 500 kg·hm-2,平作,条播,行距20 cm,以后每季将所收获秸秆全部归还小区。副区为施氮量,设纯氮75、150、225和300 kg·hm-24个施氮水平(分别用N1~N4表示)。
试验设3次重复,小区面积20 m2,供试冬小麦品种为兰天26号。试验地前茬为小麦,每处理施P2O5120 kg·hm-2。CT、CTI、PM下肥料在翻耕前一次性施入,NTS下采用免耕覆盖施肥播种机一次性完成施肥和播种。其他管理均参考当地栽培实践。试验于2014年10月8日播种,2015年7月5日收获。
1.3 试验测定指标和方法
开花期在各取样区选择表现一致、同天开花的穗挂牌标记,以后每10 d取样1次。取样时,每次各处理随机选取提前挂牌标记的样品10株,测定株高、穗长后按叶片、茎秆、叶鞘、颖壳、籽粒进行分类,并称量各器官鲜重。然后将分类的样品在105 ℃下杀青,70 ℃下烘干,测定干重、穗粒数,并计算干物质积累量。其中株高在样品分类前测定,其他指标在分类后测定。将测完干重的样品用植物样粉碎机粉碎,用于测定可溶性糖和蔗糖含量。各器官(叶片、茎秆、叶鞘、颖壳、籽粒)采用蒽酮比色法[14]测定可溶性糖和蔗糖含量。另外,在开花期和成熟期取100 cm×60 cm样方,测定叶面积[19]和穗数,同时计算开花后干物质转运量、转运效率及其对籽粒产量的贡献率:
干物质转运量=开花期地上部干物质积累量-成熟期地上部营养器官干物质积累量;
干物质转运效率=干物质转运量/开花期地上部分干物质积累量×100% ;
干物质转运对籽粒产量的贡献率=干物质转运量/成熟期籽粒产量×100%;
开花后光合积累量=成熟期籽粒产量-干物质转运量;
开花后光合积累对籽粒产量的贡献率=开花后光合积累量/成熟期籽粒产量×100%。
1.4 数据处理
用SPSS 19.0进行方差分析,用LSD法进行处理间多重比较,变量间相关分析采用Person相关系数法。用Origin 9.0作图。
2 结果与分析
2.1 耕作方式和施氮量对旱地冬小麦籽粒产量及相关性状的影响
2.1.1 耕作方式和施氮量对籽粒产量的影响
耕作方式、施氮量对旱地冬小麦籽粒产量均有极显著影响,且二者存在显著的互作效应(表1)。不同耕作方式的平均产量表现为PM>CTI>CT>NTS。其中,PM的平均产量为5 970.9 kg·hm-2,比CT高14.50%,NTS比CT减产16.74%。不同施氮量的平均产量表现为N4>N2>N3>N1,但N2、N3、N4间差异不显著。N4的平均产量达到5 477.1 kg·hm-2,比N1高14.83%。CT、CTI、NTS下,施氮的增产作用显著,但PM下不显著(表1)。CT和NTS下N4处理籽粒产量最高;CTI和PM下N2处理籽粒产量最高。在所有处理中,PMN2处理的籽粒产量最高,达到6 225.0 kg·hm-2,比最低的NTSN1高70.55%。
2.1.2 耕作方式和施氮量对产量构成的影响
耕作方式对穗粒数、穗长、千粒重、有效穗数均有显著或极显著影响(表1)。PM显著增加了冬小麦穗粒数和穗长,比最低的CTI分别增加22.17%和10.04%。NTS显著降低了千粒重和有效穗数,但其穗长和穗粒数均较高。与CTI相比,NTS的千粒重和有效穗数分别降低7.15%和21.03%。CTI的穗粒数和穗长均最低,千粒重和有效穗数最高。施氮量对穗长、穗粒数、千粒重、有效穗数的影响在不同耕作方式下存在差异(表1)。在CT下,施氮增加穗长和有效穗数,但对穗粒数无显著影响;在CTI和NTS下,施氮增加有效穗数,但对穗长、穗粒数、千粒重均无显著影响;在PM下,施氮增加穗长、穗粒数、有效穗数,但降低了千粒重。
2.1.3 耕作方式和施氮量对株高和叶面积的影响
耕作方式和施氮量对旱地冬小麦株高均有极显著影响(表1)。不同耕作方式间,PM的旱地冬小麦平均株高最高,其次为CTI和CT,最低的为NTS。除PM外,在CT、CTI、NTS下施氮显著增加了株高,N4的平均株高分别比N1增加10.40%、11.95%、12.03%。耕作方式对冬小麦开花期旗叶、倒二叶、倒三叶和总叶面积均有极显著影响。PM显著增加了不同叶位叶面积和总叶面积(表1),其中旗叶面积平均值比CT、CTI、NTS分别增加30.09%、35.99%、23.21%,总叶面积分别增加30.55%、33.43%、28.60%。施氮显著增加了不同叶位叶面积和总叶面积(表1),N2、N3、N4的总叶面积平均值分别比N1增加18.10%、28.91%、33.95%。施氮对叶面积的效应在耕作方式之间存在差异(表1)。在CT、CTI、和NTS下,叶面积随施氮量的增加而相应增加;PM下旗叶、倒二叶面积随施氮量的增加呈先增后减趋势,表现为施氮量达到高氮(N4)条件时叶面积反而降低。
2.2 耕作方式和施氮量对开花后干物质积累和转运的影响
2.2.1 耕作方式和施氮量对开花后干物质积累的影响
耕作方式和施氮量对开花后小麦植株干物质积累均有显著的影响,但影响程度因器官而异(图1)。PM显著增加了开花后不同时期茎秆、叶片、叶鞘和颖壳干物质积累量及穗粒重,但对单粒重影响不显著。不同时期茎秆、叶片、颖壳干物质积累量均随施氮量的增加而增加(图2)。施氮对粒重的影响在不同耕作方式之间存在差异,施氮增加了CT、CTI、PM下的穗粒重,降低了PM下的单粒重,对CT、CTI、NTS下的单粒重影响不显著。
2.2.2 耕作方式和施氮量对开花后干物质转运的影响
从表2可以看出,耕作方式显著改变开花后叶片、颖壳和总干物质的转运量。CT、CTI、PM、NTS下开花后叶片干物质转运量分别为839.2、839.5、1 258.5、724.2 kg·hm-2;颖壳分别为263.8、373.9、573.1、253.1 kg·hm-2;总干物质转运量分别为3 100.3、3 487.6、4 144.3、2 625.1 kg·hm-2。PM下开花后的干物质转运量显著高于其他耕作方式。施氮显著增加了开花后茎秆、叶片、颖壳和总干物质的转运量,但对叶鞘无显著影响。与N1相比,N4下开花后茎秆、叶片、颖壳和总干物质的转运量分别增加了25.91%、99.28%、61.06%和31.81%。
耕作方式对开花后叶片和颖壳的干物质转运效率有显著影响,而对茎秆、叶鞘和总干物质转运效率影响不显著(表2)。CT、CTI、PM、NTS下开花后叶片的干物质转运效率分别为58.00%、56.37%、64.14%、57.95%;颖壳分别为16.69%、21.06%、26.90%、15.87%。PM下开花后叶片、颖壳的干物质转运效率显著高于其他耕作方式。施氮显著增加了开花后叶片的干物质转运效率,而对其他器官和总干物质的转运效率无显著影响(表2)。与N1相比,N4下开花后叶片的干物质转运效率增加21.52%。
耕作方式显著影响开花后叶片和颖壳总干物质转运对籽粒产量的贡献率,但对茎秆、叶鞘和总干物质转运对籽粒产量的贡献率无显著影响(表3)。CT、CTI、PM、NTS 下叶片干物质转运的贡献率分别为13.20%、12.16%、17.17%、12.99%;颖壳分别为4.21%、5.60%、7.74%、4.57%。PM下叶片、颖壳和其总干物质转运的贡献率显著高于其他耕作方式。施氮显著增加了叶片、颖壳以及总干物质转运对籽粒的贡献率,但对茎秆和叶鞘的影响不显著(表3)。N3下叶片、颖壳和总干物质转运对籽粒贡献率最高,分别比N1高59.62%、34.50%、23.72%。耕作方式和施氮量互作效应不显著。
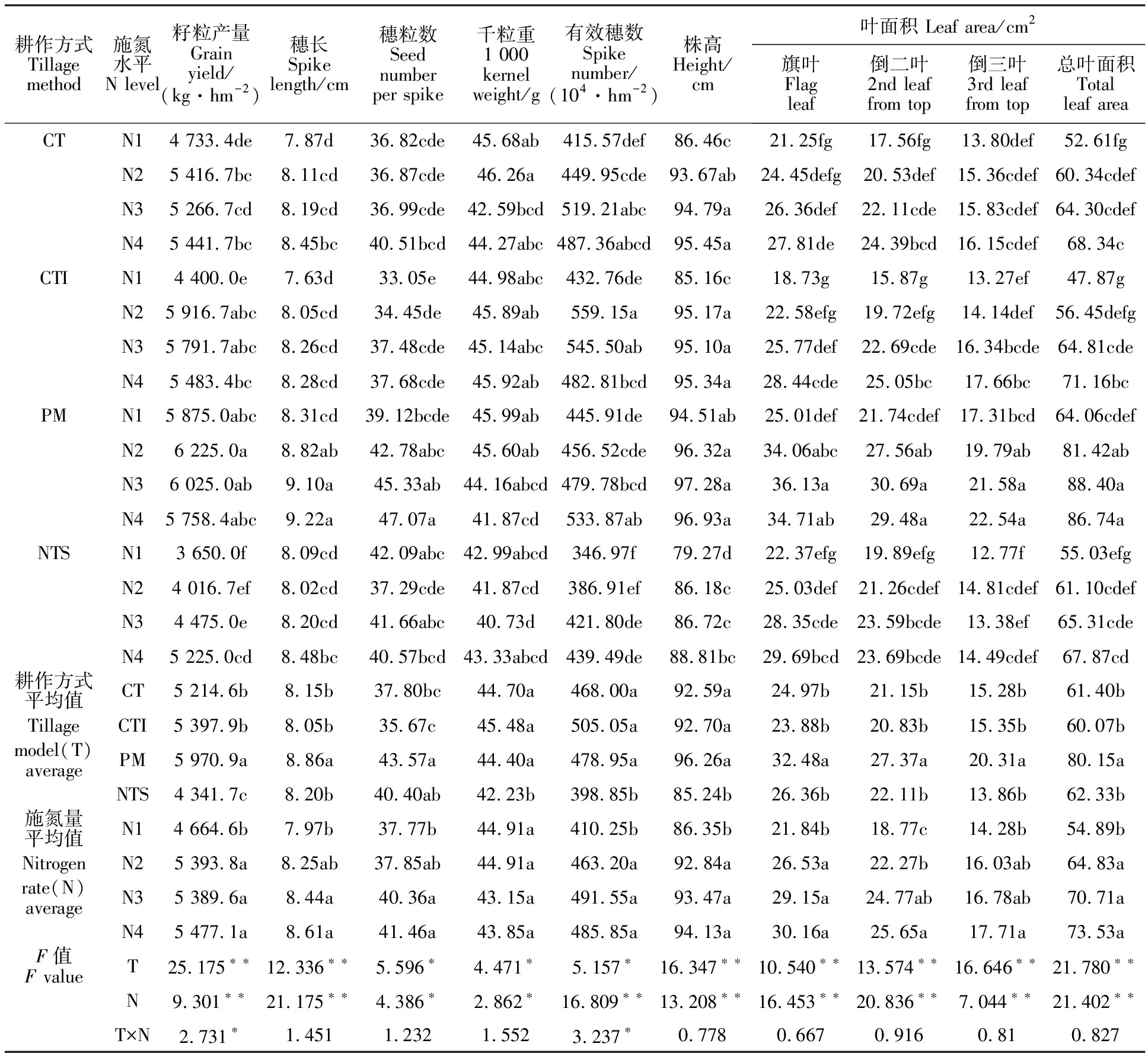
表1 耕作方式和施氮量对旱地冬小麦籽粒产量及其相关性状的影响Table 1 Effect of tillage methods and nitrogen application rate on grain yield and yield-related traits of winter wheat planted in dryland
同列数据后字母不同表示处理间差异在0.05水平上显著。*:P<0.05;**:P<0.01。下同。
Different letters within same column mean significant difference among the treatments at 0.05 level. *:P<0.05; **:P<0.01. The same in table 2 and 3.
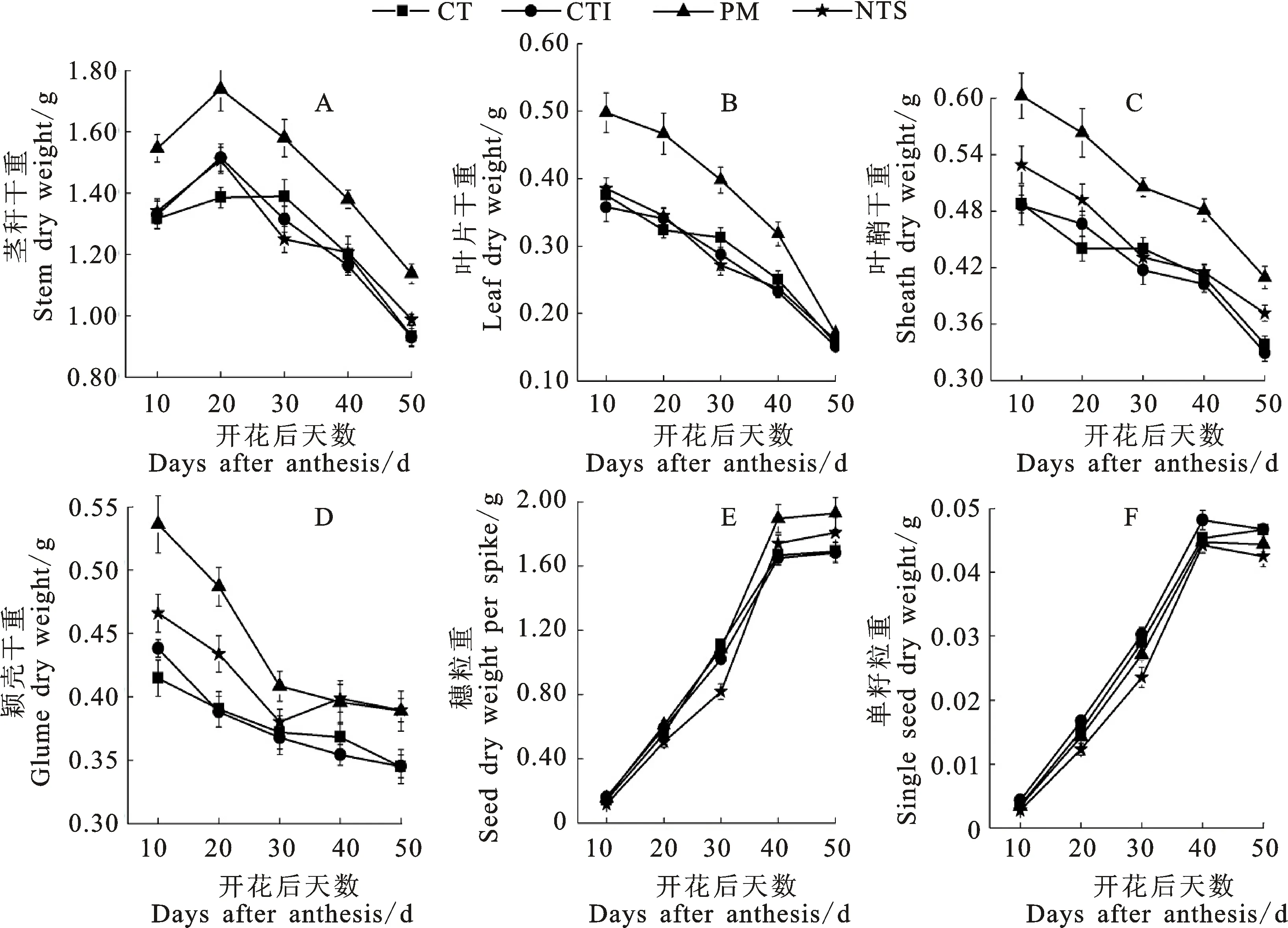
图1 耕作方式对开花后植株干物质积累和籽粒灌浆动态的影响Fig.1 Effect of tillage methods on the post-anthesis dynamic of dry matter accumulation and grain-filling
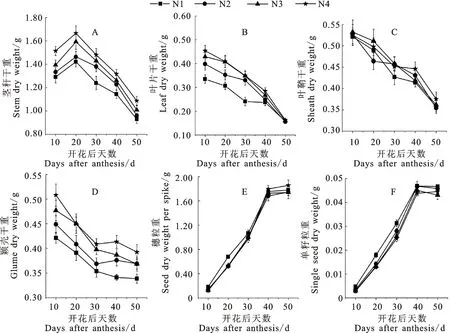
图2 施氮量对开花后植株干物质积累和籽粒灌浆动态的影响Fig.2 Effect of nitrogen application rate on the post-anthesis dynamic of dry matter accumulation and grain-filling
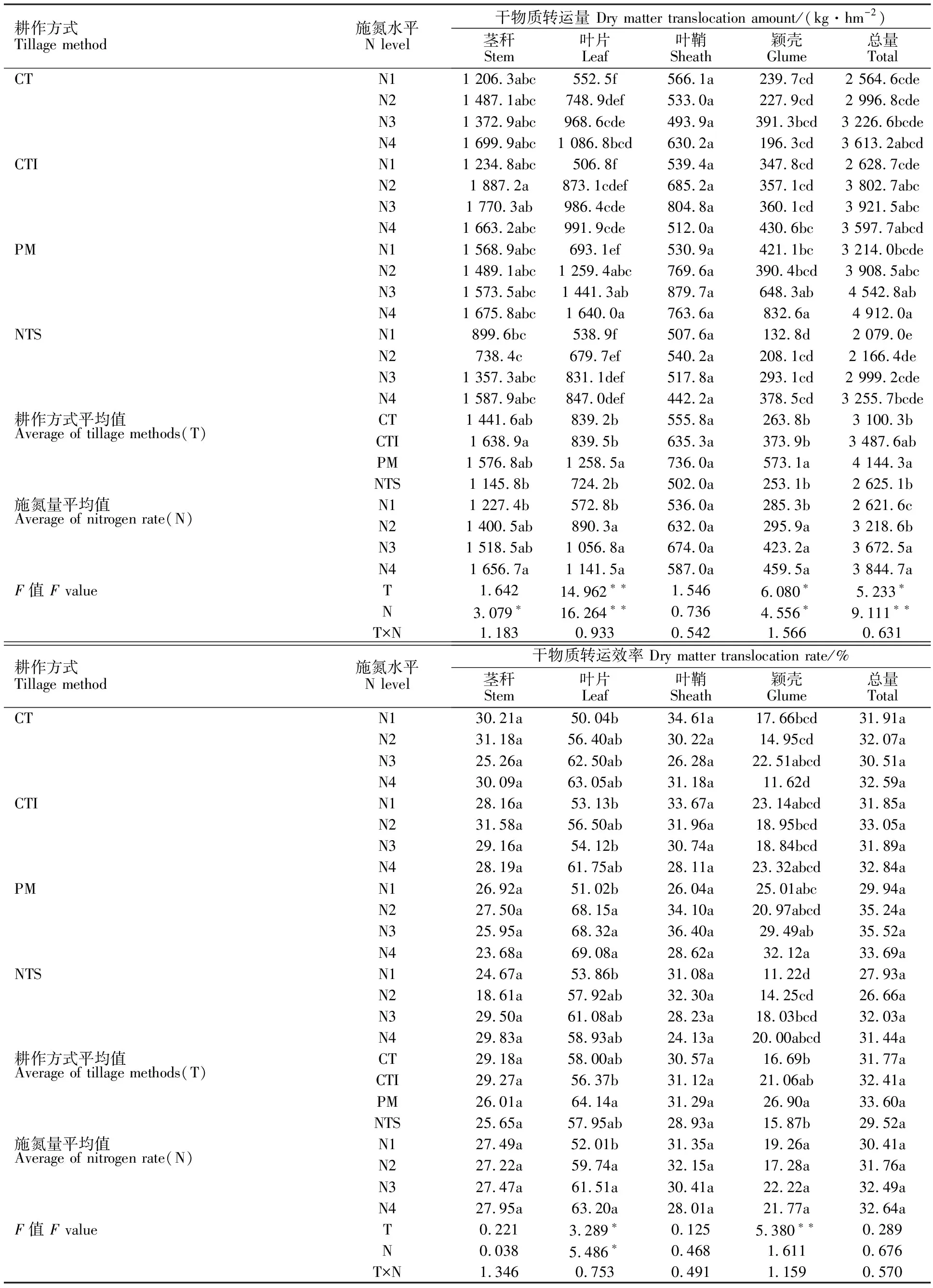
表2 耕作方式和施氮量对旱地冬小麦各器官干物质转运量和转运效率的影响Table 2 Effect of tillage methods and nitrogen application rate on the amount of dry matter translocation and dry matter remobilization efficiency
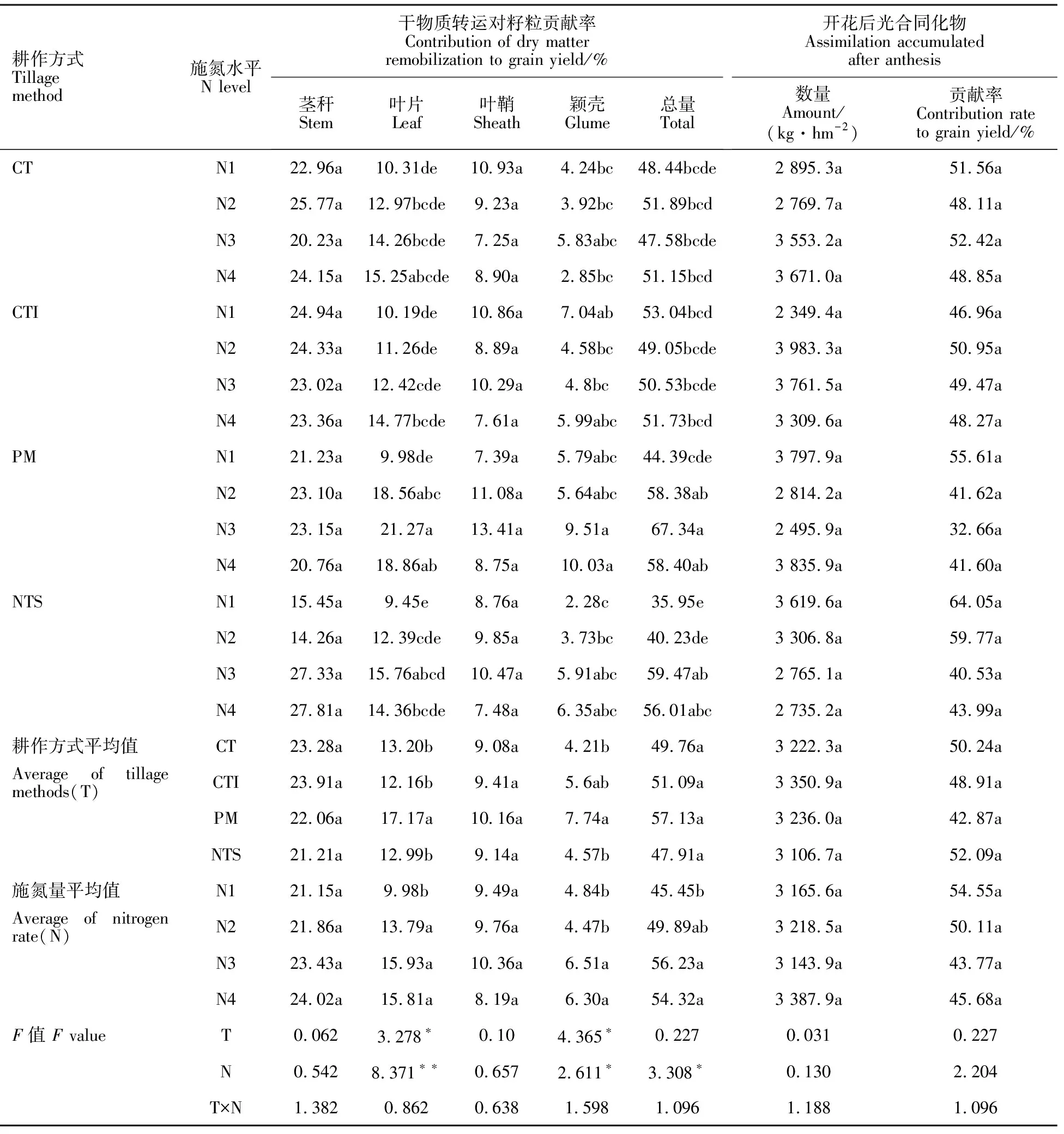
表3 耕作方式和施氮量对旱地冬小麦各器官干物质转运对籽粒贡献率的影响Table 3 Effect of tillage methods and nitrogen application rate on the contribution of dry matter remobilization to seed yield
2.3 耕作方式和施氮量对开花后可溶性糖和蔗糖积累动态的影响
从图3可以看出,耕作方式对小麦开花后各器官可溶性糖和蔗糖含量有显著影响。与CT相比,NTS下各器官的可溶性糖含量总体上增加,但因生育时期的不同而异,茎秆主要表现在开花后20~30 d,叶片和颖壳主要表现在开花后10~40 d,叶鞘主要表现在开花后20~40 d,籽粒主要表现在开花后10~30 d。PM显著提高了茎秆和叶片开花后40 d、叶鞘花后50 d的蔗糖含量。施氮显著降低了开花后茎秆和叶片开花后0~30 d的可溶性糖和蔗糖含量,但显著增加了籽粒开花后20 d的可溶性糖含量(图4)。耕作方式和施氮量互作效应不显著。
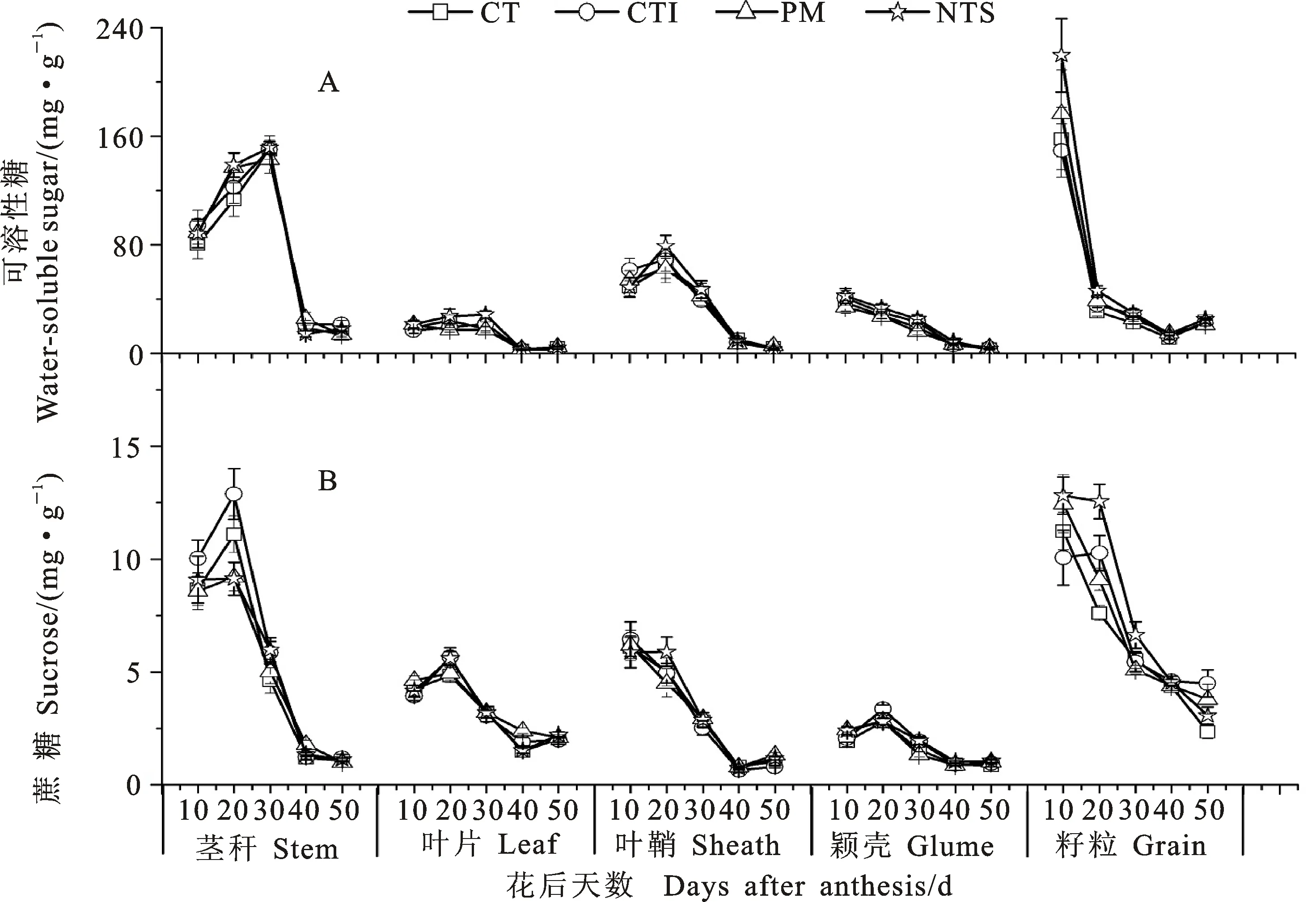
图3 耕作方式对小麦开花后各器官可溶性糖、蔗糖积积累动态的影响Fig.3 Effect of tillage methods on the post-anthesis dynamics of water-soluble sugar and sucrose in various organs
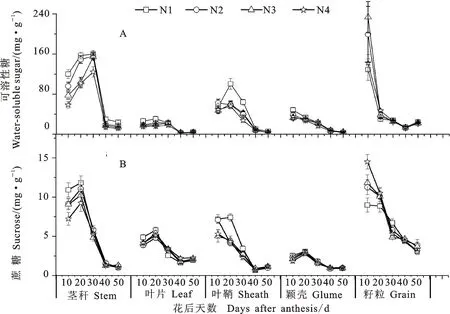
图4 施氮量对小麦开花后各器官可溶性糖、蔗糖积积累动态的影响Fig.4 Effect of nitrogen application rate on the post-anthesis dynamics of water-soluble sugar and sucrose in various organs
3 讨 论
3.1 耕作方式和施氮量对旱地冬小麦籽粒产量的影响
土壤水肥条件的改变可显著调控作物的干物质积累与分配[20]。耕作方式通过影响土壤水、肥、气、热,进而影响作物产量。覆膜能增加春小麦叶面积指数,延长了灌浆时间,增加了穗粒数、小穗数和穗数,进而提高产量[21-22]。同时,覆膜能促进冬小麦生长过程中干物质的积累及其向穗部的转运,从而提高生物量和籽粒产量;而覆盖秸秆虽能显著增加生长后期干物质积累量,但其向穗部的转运明显低于常规栽培[23]。秸秆还田显著提高了冬小麦生物量及花后干物质积累的比例,有利于粒重形成,从而获得较高籽粒产量[24]。本研究表明,4种耕作方式中,PM增产幅度最大,较CT增产14.50%。其原因主要是PM能增加旱地冬小麦株高和叶面积,使开花后“源”器官较大,进而改善开花后物质积累与分配,增加穗长、穗粒数、有效穗数,提高籽粒产量[14,22-23]。此外,PM在显著增加旱地冬小麦开花后茎秆、叶片、叶鞘、颖壳等器官干物质积累量的同时,也提高了其开花后干物质转运量、转运效率和对籽粒产量的贡献率。CTI的产量仅次于PM,比CT高3.52%,其增产主要是千粒重显著提高。NTS的株高、千粒重、有效穗数、干物质积累量和转运量均处于较低水平,因而产量最低,比CT低16.74%。本研究的这些结果与Li等[21]、Niu等[22]和李 华等[23]的研究相似。本研究中,旱地冬小麦籽粒产量随施氮量的增加而显著提高。施氮增产的原因主要是促进植株生长发育(株高、叶面积、穗长、穗粒数等),增加开花前干物质积累和开花后干物质转运,提高穗粒数、粒重、有效穗数。本研究同时也表明,氮肥不是越多越好,株高、叶面积等指标均出现边际效益递减的趋势,这一结论与Tekalign等[25]研究相似,这可能是水肥条件不适宜导致作物源/库比例不协调,导致最终不利于作物产量持续较大幅度的提高。
3.2 耕作方式和施氮量对小麦干物质转运的影响
小麦产量主要来源于花前积累的干物质在花后的再分配和花后光合同化物的积累,前者对籽粒产量的贡献率为7%~57%,特别是当灌浆期遇到逆境时作用更大[16]。干物质积累和转运显著受品种特性和农业措施的影响,覆盖(地膜和秸秆)和免耕由于改变了土壤水肥气热等微环境,从而改变了干物质的积累和转运[23,26-27]。本研究表明,与CT比较,PM显著增加了开花前干物质积累量、转运量、转运效率及其对籽粒产量的贡献率,尤其在叶片和颖壳上表现明显,这一结论与前人研究相似[23,26],主要原因是地膜覆盖在促进光合作用和干物质积累的同时,改变了小麦植株的生育进程,加速开花前期干物质积累和后期干物质的转运[23]。通常情况下,适量增加氮肥可提高叶片光合生产能力,促进营养器官建成,提高开花前后临时贮存光合产物总量,促进开花后营养器官贮存产物向籽粒的转运,提高粒重,增加产量[27]。本研究结果表明,干物质转运量和转运效率均随施氮量的增加而增加,但干物质转运对籽粒产量的贡献率在N3下达到最大,这主要是因为过量的氮肥会延长生长后期小麦叶片对蛋白质的合成作用,降低和延缓了营养体积累的干物质向籽粒的转运,最终降低籽粒产量[28]。在地膜覆盖条件下N4比N3有更低的千粒重、叶面积和籽粒产量,也进一步证明了这一结论。茎秆是花后同化物转运的主要贮存器官,其对籽粒产量的贡献率在正常情况下为10%~62%,胁迫条件下为40%~100%,旗叶光合同化物的49%会被转运到穗部[29]。本研究中,干物质转运对籽粒产量贡献率最高的器官也是茎秆,干物质转运效率最高的是叶片。另外,穗(颖壳、穗轴、芒等)跟籽粒距离较短,是花后最重要的光合器官[30]。穗部光合产物的80%被转运到籽粒中,对产量的贡献率占到10%~76%[31]。本研究得到的穗部(颖壳)干物质转运效率和对籽粒产量的贡献率分别为20.13%和5.53%,低于前人研究结果,主要原因是本研究仅仅考虑了开花前穗部同化物在灌浆期向籽粒的转运,而对开花后穗部光合同化及其对籽粒的贡献没有详细研究,并且穗部光合的潜能主要是在腊熟期后得到充分发挥[31],因此所得到的结论不能充分说明穗部光合对籽粒的贡献,还需做进一步的研究。
3.3 耕作方式和施氮量对小麦植株糖含量的影响
可溶性总糖不仅是作物的主要光合产物,也是作物体内碳水化合物转化、贮藏和再利用的主要形式,能反映叶源端的同化物供应能力和库源端籽粒对同化物的转化、利用能力,因此与作物产量密切相关[17,32]。研究表明,作物的产量与开花期植株中可溶性糖含量呈正比,尤其是与茎秆可溶性糖含量相关性更高[12-15]。本研究表明,不同耕作方式(PM、CTI)与施氮量的组合增加了花后叶面积、株高,提高花后营养器官干物质积累及转运贡献,但降低各营养器官可溶性糖和蔗糖含量。植株矮化可增加可溶性糖的含量[13-14]。本研究也证明了这一结论,不同耕作方式下冬小麦株高表现为PM>CTI>CT>NTS;不同施氮量下,株高随施氮量的增加而增加;而各器官可溶性糖含量与株高的排序刚好相反,即不同耕作模式下适度的施氮处理能促进作物体内可溶性糖的合成和积累,而在不合适的比例条件下,作物体内可溶性糖合成和积累甚至受到抑制[29]。也有研究表明,小麦茎部氮积累与可溶性碳水化合物呈显著负相关[14,33-34]。抽穗期施用氮肥会引起小麦茎秆可溶性糖含量下降[33]。因此,在低氮条件下作物往往有高的可溶性糖积累。其可能原因是充足的氮肥供应促进了植物体内氮素合成代谢,这个过程需要消耗更多的可溶性碳水化合物来提供能量,从而表现为高氮条件下植物体可溶性碳水化合物积累减少。本研究中,施氮显著降低了可溶性糖在茎秆、叶片、叶鞘、颖壳等器官的积累,相反显著增加了花后20 d籽粒的可溶性糖含量。这说明不同器官的糖分对施氮的响应与生育时期有关。
猜你喜欢
作物学报(2022年9期)2022-07-18
农业技术与装备(2022年3期)2022-05-31
山东农机化(2022年1期)2022-03-05
农业科技通讯(2022年2期)2022-02-28
腐植酸(2021年2期)2021-12-04
中国土壤与肥料(2021年5期)2021-12-02
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
养殖与饲料(2021年4期)2021-04-17
河南农业·综合版(2020年10期)2020-10-20
河南农业(2020年10期)2020-10-15
