
胶州湾大型底栖动物的次级生产力
2019-01-19 01:51刘旭东于建钊张晓红崔文连汪进生孙立娥王艳玲贾胜华
中国环境监测 2018年6期
刘旭东,于建钊,张晓红,崔文连,汪进生,孙立娥,王艳玲,贾胜华
1.青岛市环境监测中心站,山东 青岛 266003 2.中国环境监测总站,国家环境保护环境监测质量控制重点实验室,北京 100012 3.浙江省河海测绘院,浙江 杭州 310008
生产力属于生态系统生态学的部分,其基本定义为“单位时间、单位面积或体积生物有机体有机物质 (或能量) 的增加量”[1-4],一般分为初级生产力和次级生产力。次级生产力指的是动物和异养微生物通过生长和繁殖而增加的生物量或储存的能量。从能量学的角度来看,次级生产力与摄食量、同化量、排泄量以及呼吸量存在着密切关系[5]。大型底栖动物是海洋环境中的一个重要的生态类群,它在海洋生态系的能流和物流中占有十分重要的地位。以下2个特点使得底栖动物生产力的研究受到关注:①与浮游生物相比,大型底栖动物由于移动能力弱,活动范围有限,不易受短时或者随机性的短暂环境变化影响,反而能够更准确地反映出所处环境的长期、宏观变化[6];②大型底栖动物生活在泥水界面交界处,取食浮游生物、底栖藻类和有机碎屑等,本身又被经济水产动物 (如鱼类) 所食,此处产生大量的物质和能量的流动,在整个底栖生态系统中的物质循环和能量流动过程中起着承上启下的作用[7]。因此研究海洋大型底栖动物的次级生产力,不仅可帮助人们了解大型底栖动物的功能变化,而且对研究生态系统的物流、能流过程和海洋生物资源的持续利用以及评价海洋环境压力也具有重要意义[8-9]。
海洋大型底栖动物群落次级生产力的估算发展于20 世纪 70 年代,采用的多是一些经典算法,如同生群法(包括减员累计法、增长累计法、瞬时增长法、Allen 曲线法)和非同生群法(体长频率法),但有些数据不易获得,估算过程复杂费力。BREY[9]根据WARWICK[10]的建议,采用对数转换的多元线性回归法,给出了Brey(1990)经验模型,将容易获得的参数(如生物量、密度)与底栖动物的次级生产力联系起来。TUMBIOLO等[11]曾运用此公式计算了包括多毛类、软体动物、甲壳类和棘皮动物在内的34个不同海域大型底栖动物的次级生产力,验证了Brey(1990)模型能够较好地拟合这些数据,但也同时指出其估计偏差与水温和水深等环境变量密切相关。中国于2001年引入此模型[12],目前国内大型底栖动物次级生产力估算基本采用Brey(1990)经验模型[13-31]。此后国外学者又提出部分大型底栖动物次级生产力估算模型[11,32-36],并在模型中添加了环境因子(如水温、水深等参数)。但这些模型或是建模数据范围较小,不一定适用于低纬度海域[33],或是采用生物寿命等不易确定的参数,难以应用到实践[32]。DOLBETH等[35]曾在2005年评估了在不同环境下几种模型的估计效果,发现Brey(2001)[34]模型是最好的选择。此外,BREY于2012年在 Brey(2001)估算模型的基础上增加了大型底栖动物食性和移动性等易观察的数据(共计20个参数),相较以前应用的线性模型提出了改进的人工神经网络模型Brey(2012)[36],使估算结果更加真实可靠。国内目前对其他模型的应用不多,Brey(2001)模型[34]仅有几篇相关研究成果[8,30,37],Tumbiolo和 Downing 模型[11]报道更少[30],关于Brey(2012)模型,笔者只查阅到一篇文献[8]。
胶州湾是一个典型的半封闭海湾,在生态类型上更是高度人为干预下的复合生态系统的典型实例,胶州湾是中国海洋生态学研究开展最早的海域之一,对于胶州湾大型底栖动物次级生产力的研究报道始于2005年,迄今笔者只查阅到4篇相关文献[13-14,24,38],文献中均采用了Brey(1990)经验模型的计算方法,通过梳理发现存在3个问题:①在其计算过程中,不论是运用逐种累计法还是位点平均计算法,采用的均是Brey(1990)经验模型的平均参数,并未考虑底栖动物类群分类,应该根据底栖动物不同的类群选择相应的参数,这样更符合Brey(1990)经验模型的计算方式,估计结果也更加准确,目前国内已有部分研究采用这种更准确的计算方式[8,27,30-31];②次级生产力可以用湿重、干重、无灰分干重、碳含量、氮含量或能量含量等来表示,但从理论上来说,按能量测定是最真实、最合理、最具有可比性的,特别是在进行生态系统的能流分析时更应如此[5],对于胶州湾次级生产力的报道,目前均是采用的去灰干重的含量来表示,未能换算到能量含量水平;③Brey(1990)经验模型是早期的研究结果,其公式简单,仅包含生物量和生物密度2个参数,并没有包含环境及底栖动物类群的功能性特点。近年来无论是研究的深度还是广度均与国外研究存在较大差距[39]。基于以上3个原因,笔者根据2015—2017年连续3年对胶州湾大型底栖动物的监测,选取Brey(1990) 经验模型[9],Brey(2001)模型[34,40]以及Brey(2012)模型[36,40]分别估算了胶州湾海域大型底栖动物群落的次级生产力,并与胶州湾的历史数据进行对比,以期为该海域大型底栖动物的功能研究及资源利用提供参考。
1 调查与分析方法
1.1 研究区域与采样点布设
研究区域范围为胶州湾海域,避开了湾口处于航道位置及湾口北部中沙礁处附近不易采集样品的区域,共布设了12个采样点(图1),于2015—2017年每年调查春、夏、秋3个航次,调查时间分别为每年的4月下旬至5月、7月下旬至8月、10月。其中,2015年夏季航次由于潮水问题,QD07及QD08站未能成功采集样品,研究总计使用3年9个航次共106个采样点的大型底栖动物采样数据。采样使用取样面积为0.05 m2的蚌式采泥器,每个采样点取样4次,用0.5 mm孔径的网筛分选样品。样品的处理、保存、计数等均按《海洋调查规范》[12]操作,为避免混淆,研究所描述的生物量如无特殊说明均为湿重生物量(WM)。
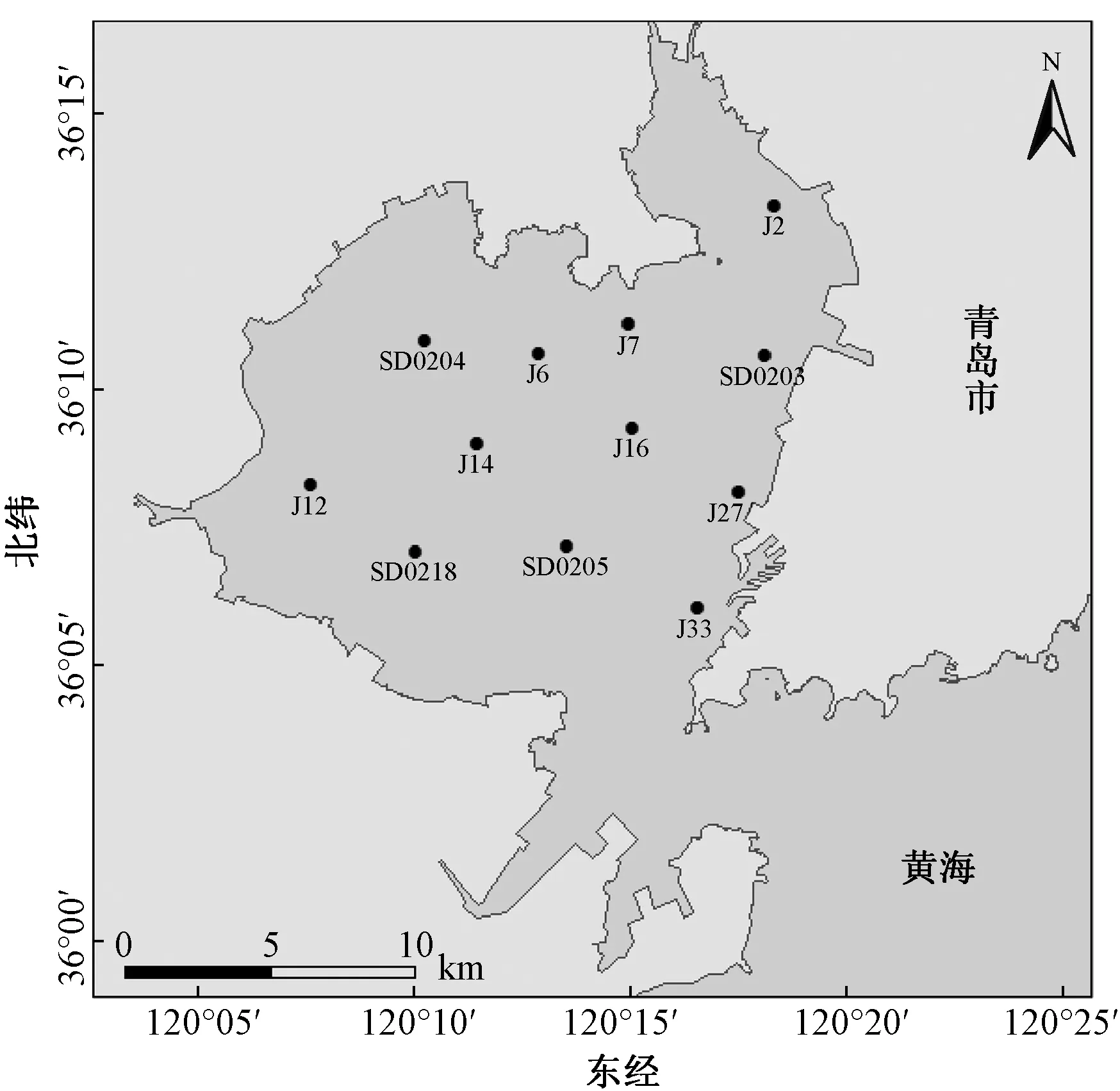
图1 采样点设置示意图Fig.1 Sketch map of sampling stations
1.2 数据处理及分析
1.2.1 Brey(1990)经验模型[9]
lgP=a+b1×lgB-b2×lgW
式中:P为年均大型底栖动物次级生产力,g/(m2·a);B为年均去灰干重生物量(湿重的9/50作为去灰干重,即湿重的1/5作为干重,干重的9/10作为去灰干重,该公式所描述的生物量按去灰干重计),g/m2;W为年均个体重量,g/ind.。不同类群的系数见表1。为了最终和另外2个模型结果对比,将该计算单位换算为kJ/(m2·a),换算比例为1 g去灰干重=0.518 g碳,1 g碳=45.7 kJ能量[41]。同时,为了和胶州湾历史调查数据对比,采用了平均(表1中其他类)参数另行计算一次。

表 1 Brey(1990)经验模型中不同类群的 3 个系数Table 1 Three coefficients for different phyla in Brey′s empirical formula
1.2.2 Brey(2001)模型[34,40]
该模型目前为4-04版本,于2004年发布,相较2001模型微调了部分参数值,具体公式为
Lg(P/B)=7.947-2.294 lg(M)-2 409.856/(T+273)+0.168/D+0.194 SubT+0.180 InEpi+0.277 MoEpi+0.174 Taxon1-0.188 Taxon2+0.33Taxon3-0.062 Habitat1+582.851 lg(M)/(T+273)
式中:P为大型底栖动物年均次级生产力,kJ/(m2·a);M为年平均个体体重能值,kJ;T为平均底层水温,℃;D为平均水深,m;B为年平均生物量对应的能值,kJ/m2;SubT:潮下带值为1,潮间带为0;InEpi:底内生物值为1,底表生物为0;MoEpi:移动生物值为1,固着生物为0;Taxon1:环节动物或甲壳动物值为1,其他类群为0;Taxon2:棘皮动物值为1,其他类群为0;Taxon3:昆虫值为1,其他类群为0;Habitat1:湖泊生境值为1,其他生境值为0。
1.2.3 Brey(2012)模型[36,40]
Lg(P/B)=a0+a1×H1+a2×H2
H1=TanH(b0+b1×lg(M)+b2×1/T+b3×lg(D)+b4×Mollusca…+b20×Exploited)
H2=TanH(c0+c1×lg(M)+c2×1/T+c3×lg(D)+c4×Mollusca…+c20×Exploited)
式中:P为大型底栖动物年均次级生产力,kJ/(m2·a);B为年平均生物量对应的能值,kJ/m2;M为年平均个体体重能值,J;T为平均底层水温,℃;D为平均水深,m;b4~b8(c4~c8)为生物类群参数,分别对应软体动物、环节动物、甲壳类、棘皮动物及昆虫类;b9~b12(c9~c12)为栖息类型及运动方式参数,分别对应是否底内生物、是否固着生物、是否只爬行、是否兼性游泳生物;b13~b15(c13~c15)为食性参数,分别对应植食、杂食及肉食动物;b16~b18(c16~c18)为栖息环境参数,分别对应湖泊、河流、海洋;b19(c19)为是否为潮间带,b20(c20)为鱼类种群,通常默认设置为0。
1.2.4 大型底栖动物能值的转换关系
根据模型对数据的要求,需将生物量湿重数据转换为能值,此项数据来自于一个关于水生生物身体成分的全球数据库[40,42]。它涵盖了体重(湿、干、无灰干重),身体成分(蛋白质、脂质、碳水化合物),元素(C、N、P)和能量含量(J)之间的比率,目前为Conversion04版本,涵盖了共计3 444种生物的5 678个数据信息。
依据胶州湾3年调查的生物种类情况, 生物类群、运动方式、食性等参数输入数据库中计算得到该次分析需要用到的转换关系(表2)。其中未能检索到螠虫动物门的多皱无吻螠(Arhynchiterugosum)及节肢动物门的日本长尾虫(Aspeudesnipponicus)的信息,由于此2种生物在调查中出现率低,且并非胶州湾内密度及生物量占据优势地位的类群,故采用与其分类及演化地位相近的星虫动物门和等足目的基准能量值来替代。

表2 大型底栖动物湿重生物量和能值的转换关系Table 2 Conversion factors of the wet weight and energy of macrozoobenthos
1.2.5 数据分析方法
事先将生产力结果进行lg(X+1)转换,并通过了正态分布和方差齐性检验。不同模型计算的生产力结果比较采用配对t检验,不同季节生产力的比较采用方差分析(ANOVA)。分析结果由SPSS 19.0完成,采样点及平面分布图由ArcGis10.2软件绘制。
2 结果
2.1 种类组成情况
历次调查共获11门19纲41目111科共191种底栖动物(附录A),其中,环节动物门多毛类出现的种类数最多,共46种,占底栖动物总种类组成的36.8%;其次为节肢动物甲壳类,共38种,占30.4%;软体动物共出现24种,占19.2%;棘皮动物出现8种,占6.4%;其余门类种数较少,合计占7.2%。3年调查中,生物种类出现数每年基本一致,各季节种类数有所不同,春夏季生物种类较多,秋季较少(图2)。

图2 调查海域的大型底栖动物组成Fig.2 Composition of each phylum of macrozoobenthos in Jiaozhou Bay
2.2 不同类群的密度及生物量
从调查数据来看,胶州湾不同调查航次的大型底栖动物类群的密度、生物量分布相差较大(表3,图3和图4),春、夏、秋季大型底栖动物的平均密度分别为595.3、505.3、277.5个/m2,平均密度为459.4个/m2,多毛类和软体动物占据密度组成主要部分,平均密度分别为200.1、166.8个/m2;春、夏、秋季平均生物量分别为1 078.5、1 011.9、394.8 g/m2,平均生物量为828.4 g/m2,软体动物是占据生物量组成的主要部分,平均生物量为786.8 g/m2。胶州湾大型底栖动物的密度和生物量均呈现春、夏季较高,秋季低的趋势,在不同季节中,密度变化与多毛类(r=0.705,P=0.034)和软体动物(r=0.794,P=0.011)的密度相关,生物量的变化则与软体动物(主要为菲律宾蛤仔)的生物量变化有关(r=0.996,P<0.001)。

表3 胶州湾大型底栖动物主要类群密度和生物量Table 3 The density and biomass of main groups of macrozoobenthos at different investigations

图3 大型底栖动物主要类群生物密度的变化分布Fig.3 The density variation of main groups of macrozoobenthos at different investigations

图4 大型底栖动物主要类群生物量的变化分布Fig.4 The biomass variation of main groups of macrozoobenthos at different investigations
2.3 3种模型的次级生产力估计
去灰干重计,Brey(1990)经验模型分类群参数估算胶州湾海域年均次级生产力为43.20 g/(m2·a),换算后为1 022.66 kJ/(m2·a);Brey(2001)模型估算胶州湾海域年均次级生产力为837.81 kJ/(m2·a);Brey(2012)模型估算胶州湾海域年均次级生产力为986.84 kJ/(m2·a)。各采样点3种估算方法的分布趋势基本一致(图5)。配对t检验分析结果显示,Brey(2012)与Brey(2001)模型估算结果并无显著性差异(t=0.397,P=0.692);Brey(2012)与Brey(1990)经验模型估算结果差异显著(t=5.561,P<0.001);Brey(2001)与Brey(1990)经验模型估算结果差异显著(t=6.067,P<0.001)。
此外,采用Brey(1990)经验模型平均参数估算胶州湾海域年均次级生产力,结果为54.71 g/(m2·a),高于按类群估算为43.20 g/(m2·a)的结果(t=9.690,P<0.001)。

图5 不同估算方法的胶州湾大型底栖动物次级生产力的变化趋势Fig.5 The change in the secondary production of macrozoobenthos among different models
3 讨论
3.1 不同海域次级生产力的比较
在进行底栖动物群落历史对比分析时,由于以往调查者的目的和侧重点有所差异, 调查采样点和监测水平不一致,调查结果与采样航次、采样点是否均匀覆盖、季节性有很大关系。该研究结果与对胶州湾次级生产力的历史研究[13-14,24,38]结果整体趋势一致,即胶州湾大型底栖动物次级生产力由湾口到湾北部的湾顶呈现升高趋势,菲律宾蛤仔是次级生产力的主要贡献者,菲律宾蛤仔资源的贡献使得胶州湾大型底栖动物的次级生产力远高于其他海域的次级生产力(表4)。
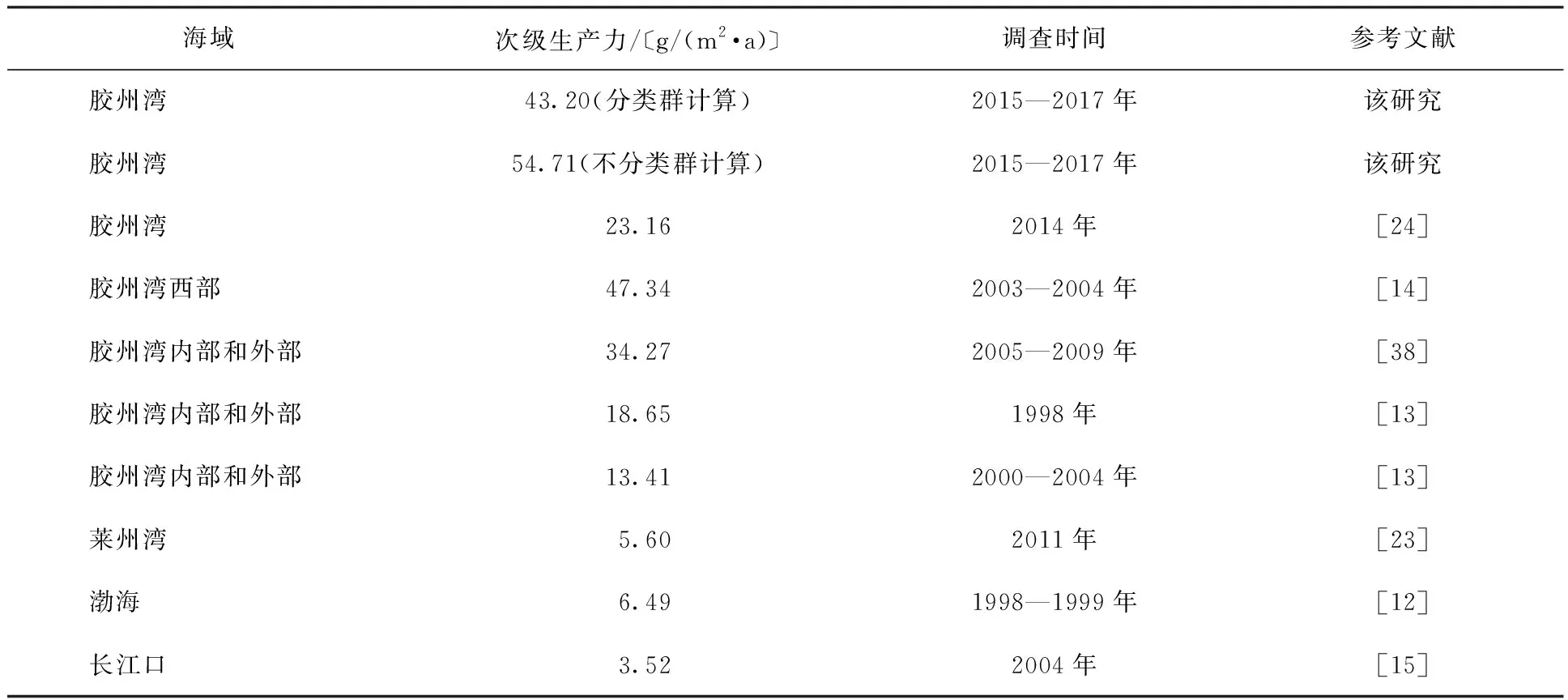
表4 胶州湾大型底栖动物平均次级生产力与其他海域的比较Table 4 The secondary production of macrozoobenthos in Jiaozhou Bay and the comparison with other seas
研究所调查的胶州湾海域大型底栖动物的次级生产力较历史资料高有以下几个原因:①调查的采样点均分布于胶州湾内,并避开了水深较深的航道区域及不易采样的中砂礁位置,与其他调查航次采样点相比,处于其高值区[13,38];②次级生产力与水温密切相关,升高温度可加快底栖动物的生长发育速度,进而提高生产力[5]。研究未能调查冬季航次,因此在估算年均生产力时可能会导致结果偏高,推荐根据实际情况采用按季节估算的结果;③菲律宾蛤仔是次级生产力的主要贡献者,该次调查中有很多采样点位于菲律宾蛤仔养殖区,可采到大量的菲律宾蛤仔(如在J6采样点,2016年春季航次中菲律宾蛤仔的密度达到1 060个/m2),因此模型的估计结果受菲律宾蛤仔资源的波动影响。
3.2 合理模型的选择
经验模型只是对群落次级生产力的估算,并非真实值,研究所采用的3个模型逐步复杂精细化,这与近年来对于次级生产力研究的发展相关。其差异主要表现在2个方面:①模型考虑的参数由2个增加到20个(如增加了底栖动物的食性、运动方式情况),同时增加了环境因子;②能值转换以前所有底栖动物均采用一个转化基准,随着研究的进一步深入,现在精细到更细的类群专属转换比例。研究显示所用的3种经验模型得到的次级生产力估算存在差异,Brey(1990)经验模型估算结果偏高。而关于Brey(1990)经验模型与Brey(2001)模型估算结果差异性显著的情况,均在国内的3篇报道[8,30,37]中出现,一般认为Brey(1990)经验模型公式中未涉及环境参数,实质上仅给出特定环境条件下群落次级生产力的平均水平,这可能使 Brey(1990)经验模型公式对不同季节和不同环境的次级生产力估算效果不理想[8,30,37]。此外在胶州湾海域中采用Brey(1990)经验模型不按类群划分计算的情况,其结果高于分类的计算结果,因为软体动物菲律宾蛤仔是胶州湾大型底栖动物次级生产力的主要贡献者,其计算系数2种方式有较大差别,在该研究中不按类群划分会导致计算结果高估26.7%。
从建模方法看,Brey(1990) 经验模型[9]与Brey(2001)模型[34,40]采用的为多元线性回归法(MLM),而Brey(2012)模型[36,40]采用人工神经网络法(ANN)。MLM对独立变量间的交互性非常敏感,而ANN可以对自变量和因变量间复杂的、非线性的、间断性的关系建模。大型底栖动物群落次级生产力估计上,MLM法优于ANN法[36]。因此,虽然3种模型估算的胶州湾大型底栖动物次级生产力的时空变化趋势基本一致,Brey(2001)模型与Brey(2012)模型的估算结果无显著性差异,该研究仍推荐采用Brey(2012)模型进行胶州湾大型底栖动物次级生产力的估计,并采用Brey(2012)模型的计算结果来讨论胶州湾大型底栖动物次级生产力的时空分布。
3.3 胶州湾大型底栖动物次级生产力的时空分布
由于存在影响生态系统的多重压力因素(如富营养化、污染、栖息地丧失、气候变化、入侵物种引进等),可以通过对种群的静态和动态分析检测到这些变化,次级生产力在实际应用中提供的信息远比单独的密度或生物量等其他静态测量方法所提供的信息多,它结合了种群生态性能的静态和动态组成部分,并不仅仅服务于能量流动和利用效率的研究[43],如图6所示,2017年春季具有较高的密度和生物量水平,但生产力却处于平均水平;2015年夏季生物量处于一般水平,但生产力却处于最高水平;2015年春季密度处于较高水平,但生产力却处于低水平。
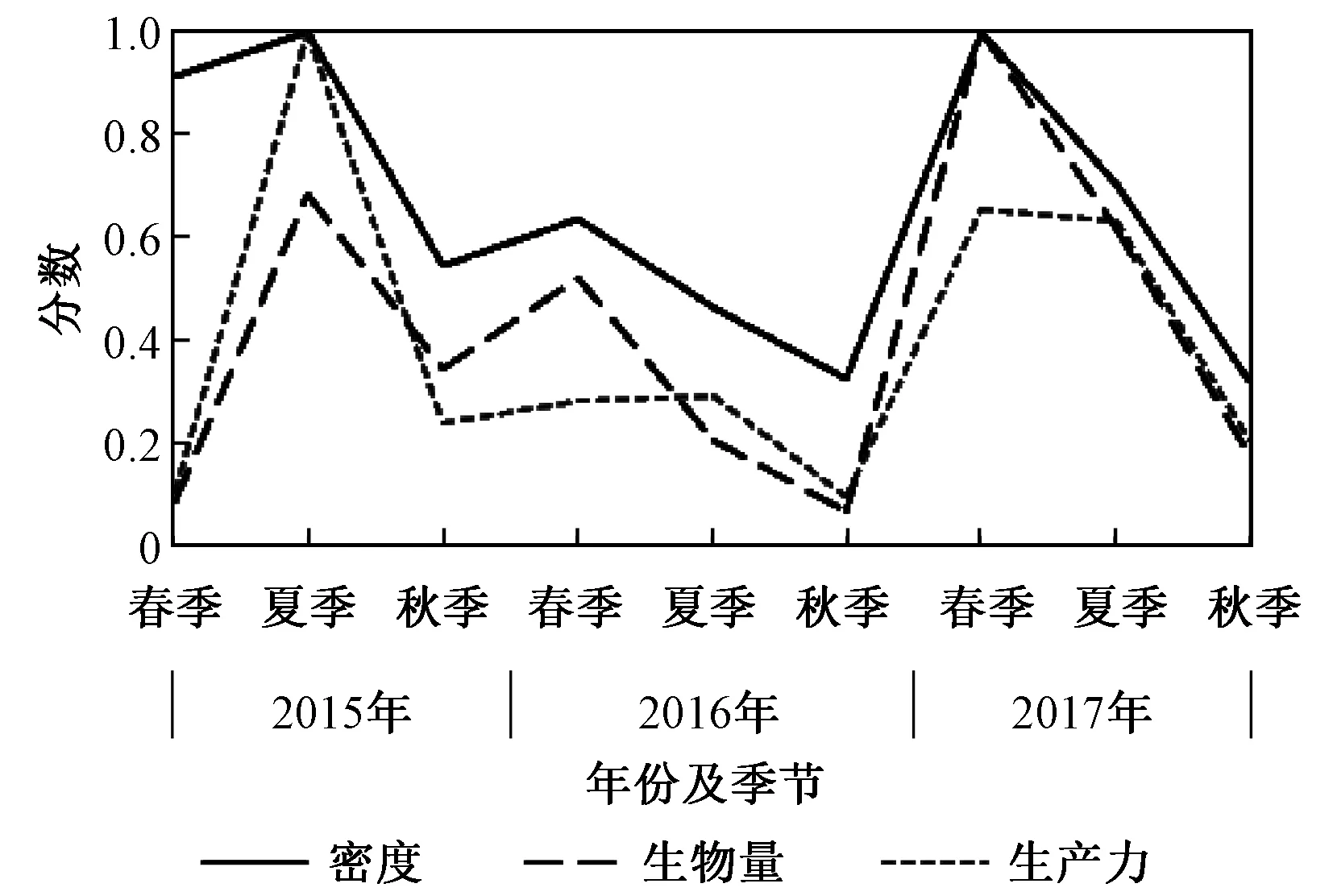
注:得分为1~0,分别代表每个参数在3年研究中的最高和最低值。图6 不同调查时期的密度、生物量及次级生产力的得分Fig.6 Scores for the secondary production, density and biomass for the macrozoobenthos of Jiaozhou Bay from 2015 to 2017
胶州湾大型底栖动物具有较高的生产力水平(一般也意味着高经济水平),但是高水平的生产力并不一定代表一个更健康的生态系统,一些影响可能导致整体底栖动物产量的增加,因为环境的变化有利于高产机会主义物种,但同时也降低了种群的复杂性(降低均匀度和丰富度)[43],正确认识胶州湾大型底栖动物的次级生产力来源对研究胶州湾海域的大型底栖动物群落生态环境状况也具有重要意义。通过该研究发现,胶州湾大型底栖动物各类群所占比例相比于历史数据并没有发生太大变化,该研究的结果是环节动物多毛类(36.8%)>节肢动物甲壳类(30.4%)>软体动物(19.2%);徐兆东[24]2014年研究结果为环节动物多毛类(41.8%)>节肢动物甲壳类(29.3%)>软体动物(20.5%);于海燕等[44]1998—2001年的研究结果为多毛类(41.3%)>节肢动物甲壳类(28.6%)>软体动物(18.3%),孙道远等[45]1991—1994年的研究结果为多毛类(39.0%)>节肢动物甲壳类(28.5%)>软体动物(20.0%)。对比发现胶州湾历次调查大型底栖动物类群顺序相同,所占比例极其接近,说明胶州湾近30年的大型底栖动物在类群组成上并没有发生明显变化,处于稳定状态。胶州湾大型底栖动物主要的次级生产力贡献来源于菲律宾蛤仔。
胶州湾海域大型底栖动物次级生产力呈现出夏季>春季>冬季的变化趋势(表5),季节间差异显著(F2,36=4.351,P=0.021),春、夏、秋季的估算结果分别为880.17、1 537.64、458.33 kJ/(m2·a)。主要是夏季水温较高,菲律宾蛤仔较活跃,从而具备较高的生物量和密度。在空间分布上(图7),高次级生产力的区域为处于湾顶中部红岛以南到向西偏移至大沽河口和红石崖附近,这也与以往研究一致[13-14,24,38]。总体来看,胶州湾大型底栖动物次级生产力具备较高的水平,且具有显著的时空变化,菲律宾蛤仔资源的变化是造成波动的最主要因素。
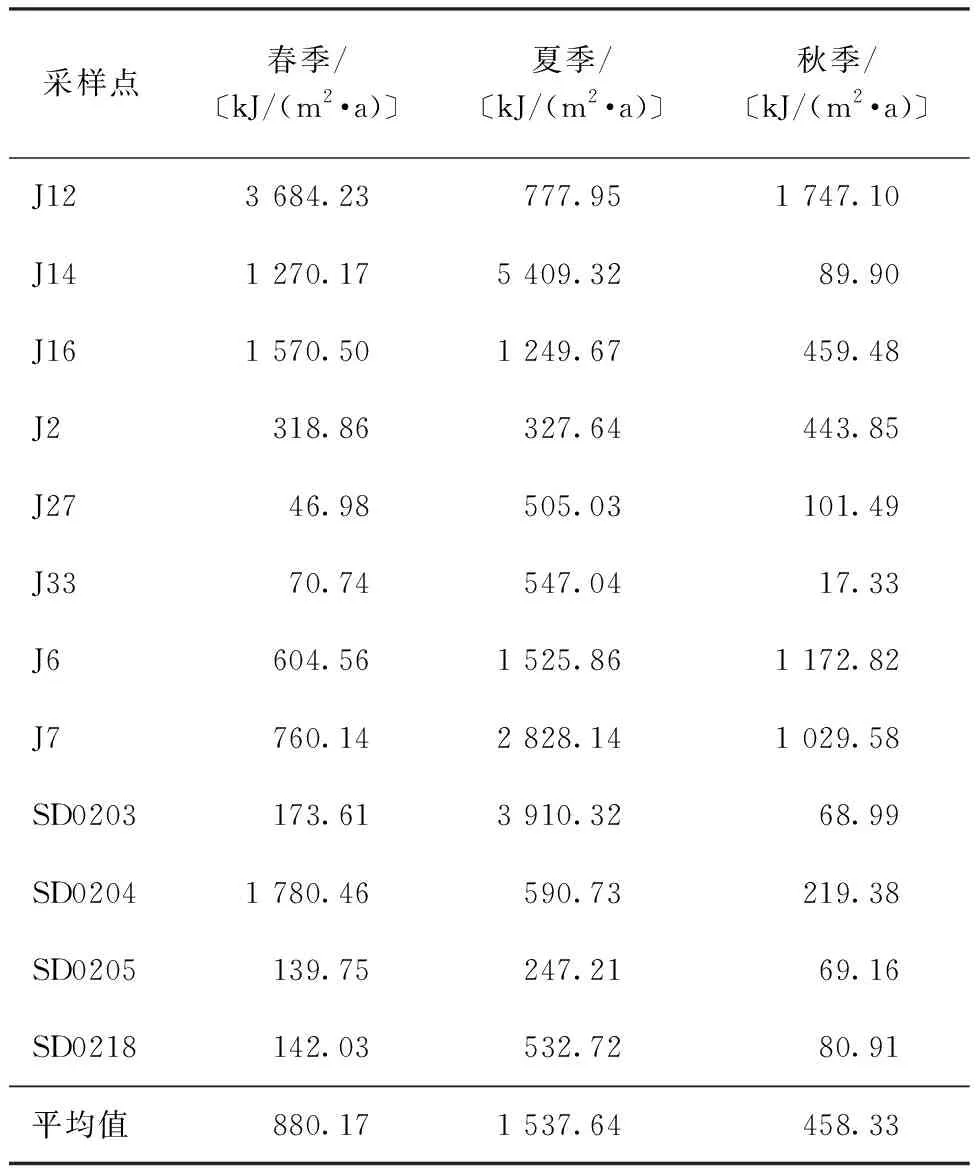
表5 不同季节胶州湾大型底栖动物的次级生产力Table 5 The secondary production of macrozoobenthos in Jiaozhou Bay at different seasons

图7 胶州湾大型底栖动物的次级生产力分布Fig.7 The secondary production distribution of macrozoobenthos in Jiaozhou Bay
4 结论
胶州湾近30年的大型底栖动物在类群组成上并没有发生明显变化,处于稳定状态。胶州湾大型底栖动物次级生产力由湾口到湾北部的湾顶呈现升高趋势,菲律宾蛤仔是次级生产力的主要贡献者,也导致胶州湾大型底栖动物的次级生产力远高于其他海域。
相较其他估算模型,推荐使用Brey(2012)模型估算胶州湾海域大型底栖动物的次级生产力,胶州湾海域春、夏、秋季的估算结果分别为880.17、1 537.64、458.33 kJ/(m2·a)。
猜你喜欢
生态学报(2021年3期)2021-03-31
绿色科技(2019年14期)2019-11-19
江苏农业科学(2019年5期)2019-09-02
陕西农业科学(2019年6期)2019-07-19
东方企业家(2018年8期)2018-08-22
红土地(2016年3期)2017-01-15
环球市场信息导报(2015年15期)2015-12-03
海峡姐妹(2015年9期)2015-02-27
中国海洋大学学报(自然科学版)(2014年6期)2014-02-28
公务员文萃(2013年5期)2013-03-11
