
不同生境下景宁玉兰灌丛萌株形态及其生殖特征
2019-01-18 05:51杜有新吴伟建刘跃钧王军峰季志利章伟民何小勇
生态学报 2018年23期
杜有新,吴伟建,刘跃钧,王军峰,季志利,章伟民,何小勇,*
1 丽水市林业科学研究院,丽水 323000 2 松阳县林业局,松阳 323400 3 丽水莲都生态林场,丽水 323000
景宁玉兰(MagnoliasinostellataP. L. Chiu et Z. H. Chen) 属浙江特有种[1- 2],为国家二级保护的濒危物种[3- 4],具有较高的科学研究和园林观赏价值[5]。景宁玉兰集中分布于浙江南部的丽水市景宁畲族自治县草鱼塘林场、松阳县牛头山地区、莲都区峰源乡和温州市乐清雁荡山,其原生环境相似,由于人类活动的干扰及自然环境变化导致的生境破碎化,现代分布格局呈分散性特征[6],主要生长于海拔900m以上的落叶阔叶林、杉木林、黄山松林、林缘沟边和灌丛中[7]。
景宁玉兰为浅根性落叶灌木,喜阴凉湿润、水分充足且土壤肥沃的生境,对环境变化较为敏感[7- 8]。在自然环境下,景宁玉兰自然结实率不到1%[9],但其萌生性强,茎秆基部具有很强的萌芽能力[2],实生苗定植后往往萌发数根萌蘖[10],根颈萌枝[11]和残桩萌枝[12]现象在原生环境下普遍存在,这也是其对资源状况及环境变化的一种适应性响应[13-15]。之前对景宁玉兰生态生物学特性[2]、种群分布[6- 7]、花粉活力[8- 9]、无性繁殖[10]及属内亲缘关系[16]等方面开展了较多研究,但未见有关不同生境下景宁玉兰萌枝形态及生殖特征方面的报道。萌生是植物抵抗不利环境的一种营养繁殖方式一,是面临较大强度干扰及实生更新受到限制情况下的一种重要自然更新方式[17],研究萌枝形态及其生殖特征可以深入了解植物在不利环境下的生长发育状况及其应对策略。本文通过调查分析不同生境下景宁玉兰萌枝形态及开花结实状况,以期了解景宁玉兰自然更新特征及其与生境的关系,为科学保护合理利用其种质资源提供参考。
1 研究地概况
本研究选取景宁玉兰代表性的3个分布点即丽水市松阳县大东坝镇灯塔村牛头山、丽水市莲都区峰源乡尤源村和葑垟村,3个分布点相距20—60km。分布点均处于山体北坡,含盖了景宁玉兰自然生长的各种生境类型,除了松阳牛头山海拔720m处生长着3丛外,景宁玉兰主要分布于海拔950—1300m之间。景宁玉兰在浙南山区的分布区位于27°30′—28°26′N,119°49′—120°05′E之间的北坡[6],土壤为山地黄壤和黄棕壤,土层厚度30—120cm,pH值4.8—5.5[2]。其中在峰源乡的自然分布区位于28°05′—28°14′N,119°40′—119°51′E之间,年均气温处于12.0—15.5℃,海拔950—1100m,年均相对湿度87%[18]。牛头山分布区处于28°15′—28°17′N,119°32′—119°35′E之间,年均气温处于11.5—16.5℃,海拔700—1285m,年均相对湿度87%。研究样地的地理信息及植被概况见表1。
2 研究方法
2.1 样地设置
于2016年2月,根据景宁玉兰自然生长的生境条件,把景宁玉兰生境分为5种类型,分别为灌丛(SH)、黄山松(Pinustaiwanensis)林(PTF)、落叶阔叶林(DBF)、杉木(Cunninghamialanceolata)林(CLF)及林缘沟边(FE),其中林缘是指树林边缘5m内区域,距离树林边缘10m视为林内。根据景宁玉兰分布状况,在每种生境上分别设置1—3个典型样地,样地面积10m × 20m,每个样地划分为8个5m × 5m的小样方,尤源村共设置了4个样地;葑垟村灌丛生境设置3个10m × 20m样地,而林缘生境的景宁玉兰居群主要分布于长为100余m的毛竹林和杉木林的林缘沟边,设置2个面积5m × 40m样地,每个样地划分为8个5m × 5m的小样方(表1)。在松阳县牛头山地区,景宁玉兰分布于海拔716—1300m之间,结合景宁玉兰种群数量,在海拔720、1100m和1250m处分别设置1、2和3个样地。海拔720m处只发现相距不到5m的3丛,直到海拔1030m后才再次出现景宁玉兰植株,另外景宁玉兰在海拔1000—1200m的落叶阔叶林中只零散分布着9丛,因此,在720m处只设置了1个样地,在落叶阔叶林生境设置2个样地,牛头山上共设置6个样地(表1)。

表1 景宁玉兰群落样地概况(均值±标准差)
SH,灌丛,Shrubberies; PTF,台湾松林,Pinustaiwanensisforest;DBF,落叶阔叶林,Deciduous broad-leaved forest;CLF,杉木林,Cunninghamialanceolataforest;FE,林缘,Forest edges
2.2 调查方法
由于灌丛年龄结构的确定受到方法上的限制,通常采用立木的大小径级代替不同的年龄结构,但至今也没有景宁玉兰径级与树龄关系的资料。景宁玉兰属世代重叠及生殖连续的多年生植物,笔者根据实际调查和相关文献[19],以景宁玉兰每丛最大萌枝作为母株,按其基径(BD)大小划分为5个年龄级,分别为I:BD<1.5cm,II:1.5≤BD<3cm,III:3≤BD<4.5cm,IV:4.5≤BD<6cm,V级:BD≥6cm。本研究样地上的景宁玉兰均为天然更新苗,根据上述标准,所有样地景宁玉兰种群年龄均处于II至IV之间。
调查统计样地内景宁玉兰株(丛)数及其冠幅,所有从母株根颈部或其地表根出萌枝均视为其萌枝,观测每丛所有萌枝的基径和高度等信息,记录伴生植物种类,同时记录样地的经纬度、海拔、坡向、坡度等生态因子及干扰情况。景宁玉兰原生地存在从母株根部萌生的根出条萌枝和机会萌枝[11]现象,在实际调查中把与母株相连的萌枝均视为母株萌枝,不计为独立的植株,野外调查中没发现萌生起源的植株。在每个样地内选取2—3丛代表性成年(III级)植株,在其冠层上部标记3根主枝,用直尺和游标卡尺测量各级枝的基径和枝长。按Strahler法确定分枝序列[20],冠层最外层的第一小枝为第一级,两个以上第一级相遇即为第二级,两个以上第二级相遇后则为第三级,依此类推。光照强度采用浙江托普仪器公司的农业环境检测仪(型号DJL- 18)测定。
于2016年2—3月,对每个样地景宁玉兰植株及每丛萌枝全部编号挂牌,于2016年和2017年的2—3月份,每丛开花的萌枝计为生殖萌枝,花期调查每根生殖萌枝花量,统计每丛花量及每个样地开花丛数。于2016年和2017年的8—9月,在景宁玉兰果实成熟期,对每个样地景宁玉兰植株,根据花期调查时每个样地上的编号植株,逐丛逐根萌枝进行结实情况调查,对采集的果实进行编号,用直尺和游标卡尺测量每个果实的长度和直径,果实开裂后记录每个果实种子粒数,统计生殖萌枝结实率和每丛结实率。
2.3 数据处理与分析
萌枝总分枝率(Overall bifurcation ratio,OBR)和逐级分枝率(Stepwise bifurcation ratio,SBR)分别采用如下公式[20]计算:总分枝率OBR=(NT-Ns)/(NT-N1),逐级分支率SBRi:(i+1)=Ni/N(i+1),式中NT为所有枝级中的枝条总和,NS为最高级枝条数,N1为第一级的枝条数,Ni和Ni+1分别为第i级和第i+1级的枝条数。生殖萌枝花量为每根生殖萌枝着花的数量,每丛生殖萌枝比例=每丛开花的萌枝数/每丛萌枝总数;生殖萌枝结实率=每根生殖萌枝果实数量/每根生殖萌枝花量总数,每丛结实率=每丛果实总数/每丛花量总数。
把松阳县牛头山的景宁玉兰居群划分为3个海拔梯度,即海拔700—1000m、1000—1200m和>1200m。分析海拔高度对景宁玉兰形态及生殖性状的影响。选取海拔950—1200m处的灌丛、落叶阔叶林、杉木林和林缘等4种生境,分析不同生境类型下景宁玉兰形态及生殖性状差异。通过单因素方差分析(ANOVA)和多重比较(LSD)检验不同植被类型和不同海拔高度间景宁玉兰萌枝形态及生殖性状的显著性差异,采用Pearson相关性分析检验各性状之间的相关性特征。所有数据均在Excel 2003和SPSS 17.0软件上进行。
3 结果与分析
3.1 不同生境景宁玉兰萌枝形态特征
景宁玉兰萌枝形态特征及分枝率在不同生境之间存在一定差异(表2)。黄山松林下的景宁玉兰密度最高,每公顷平均达732丛,显著高于落叶阔叶林和杉木林(P<0.05),而且落叶阔叶林下景宁玉兰萌枝基径显著小于其他林分(P<0.05)。在分枝特征方面,黄山松和落叶阔叶林下景宁玉兰一级枝基径显著小于其他3种生境(P<0.05),黄山松林下景宁玉兰一级枝长度即年生长量显著低于其他4种生境(P<0.05)。灌丛中景宁玉兰的总分枝率显著高于杉木林落叶阔叶林和黄山松林(P<0.05),灌丛和林缘生境上的景宁玉兰逐步分枝率均处于较高水平,也许光照条件不受限制,高分枝率有利于拓展更大的生存空间。

表2 不同生境下景宁玉兰形态特征 (均值±标准差)
同列中不同小写字母者表示差异显著水平(P< 0.05)
3.2 不同生境景宁玉兰生殖特征
不同生境下景宁玉兰平均生殖萌枝比例及生殖萌枝开花数量存在差异(表3)。其中黄山松林、落叶阔叶林和林缘的景宁玉兰生殖萌枝花量显著低于杉木林(P<0.05),灌丛与其他生境没有差异。黄山松林下景宁玉兰生殖萌枝比例显著低于灌丛、杉木林和林缘(P<0.05),落叶阔叶林下景宁玉兰生殖萌枝比例显著低于林缘。景宁玉兰萌枝生殖性状除落叶阔叶林外,其他4种生境上的景宁玉兰生殖萌枝结实率没有差异。黄山松林下的景宁玉兰果实大小、单果种子数及每丛结实率最高(P<0.05),落叶阔叶林下景宁玉兰没有结实植株,其他3种生境没有差异。景宁玉兰生殖特征也许与其受到的光照资源及花期气温密切相关,有待深入研究。
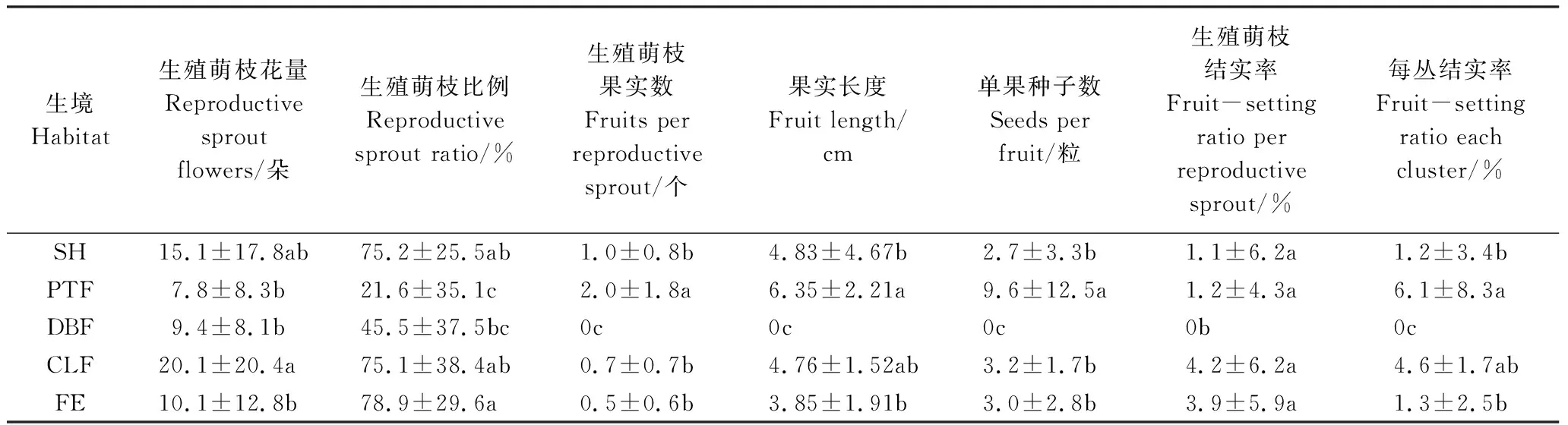
表3 不同生境下景宁玉兰生殖特征(均值±标准差)
3.3 萌枝性状相关性分析
通过萌枝基径与高度之间及其与萌枝花量的相关分析表明,萌枝基径与高度具有极显著正相关关系(P<0.01),萌枝基径(P<0.01)和高度(P<0.01)与萌枝花量均有极显著正相关关系(图1)。每丛最大萌枝基径与高度之间具极显著正相关关系(P<0.01),最大萌枝基径与每丛萌枝数具有显著正相关关系(P<0.05),而最大萌枝高度与每丛萌枝数没有相关关系(图1)。通过生殖萌枝和每丛花量及其与结实率之间相关分析表明,生殖萌枝花量与萌枝结实率没有关系(P>0.05),每丛花量与每丛结实率之间也没有关系(P>0.05)。结果说明萌枝大小与其高度密切相关,并影响着景宁玉兰的开花数量,但每丛开花数量与结实状况没有显著相关关系。

图1 萌枝形态和生殖性状的相关性Fig.1 Relationship between morphological and reproductive characters of sprouts
3.4 萌枝性状的变异性分析
生境类型和海拔高度显著影响着景宁玉兰萌枝的基径、高度和一级枝长度(P<0.05),海拔对一级枝粗度和长度具有显著影响(P<0.05),但生境类型和海拔高度对每丛萌枝数及分枝率均没有影响(表4)。生境类型和海拔高度对生殖萌枝花量、生殖萌枝比率及生殖萌枝结实率具有显著影响(P<0.05)。说明景宁玉兰萌枝形态及其生殖生物学性状与植株所处的生境条件密切相关。

表4 生境类型和海拔对景宁玉兰萌枝形态及生殖萌枝特征的影响
*和**分别表示显著性差异(P<0.05)和极显著性差异(P<0.01)
4 讨论
植物的萌生能力受内在生物学特性及微生境、资源水平和干扰状况等外部因子的综合影响[21]。景宁玉兰属萌生能力较强的密集型生长类型[10],以根颈萌生类型为主[2, 14]。萌生能力与母株的大小存在一定的相关关系,但萌枝数量与母株高度没有相关关系(图1 E—1F),这与萌生能力很强的亚热带乔木树种米心水青冈(Fagusengleriana)有点不同,米心水青冈的萌生能力与母株的大小以及萌枝数量与母株高度均呈正相关关系[22]。与温带落叶树种连香树(Cercidiphyllumjaponicum)也不同,连香树萌枝数与立地条件和根系暴露程度有关[23],而景宁玉兰植株萌枝数量在不同生境之间也没有差异,如在人类活动较为频繁的尤源村灌丛和没有干扰的牛头山黄山松林中,景宁玉兰每丛的萌枝数量没有差异,但不同生境之间的萌枝大小及高度存在差异(表2),说明景宁玉兰的萌枝能力很可能主要由其生物学特性控制[11],而萌枝大小及高度差异可能是萌枝间以及个体间光照及土壤养分资源的竞争结果[14,24],有待深入研究。
萌枝形态特征能较好地反映植物对资源与环境变化的适应状况[20]。虽然适度低温和阴湿环境是玉兰亚属较为理想的生存环境[25],但景宁玉兰在不同生境上,因为所处的微生境和面临的限制因素各不相同,分枝构型在不同生境上的表现有较大差异。对林下植物而言,有机物质积累与其接受的光资源密切相关,光资源条件是影响植物萌蘖和增粗的主要因素[15],如黄山松和阔叶林下生境,由于上层乔木树种遮蔽,具有林下光照较少、种内种间竞争较为激烈,景宁玉兰可能通过低分枝率(表2)以增加有效光合作用面积。海拔通过环境条件的变化间接影响着植物生长[26],中低海拔的阔叶林和杉木林下的景宁玉兰一级枝粗度和长度显著高于黄山松林下,可能与土壤有机质和碱解氮含量有关[27]。植物群落及土壤环境对景宁玉兰生长及枝型特征均有影响,虽然不同生境上植被群落性质不同[6]和缺乏完整的土壤环境数据,但由于林缘生境的灌木覆盖度较低和灌从生境上没有上层林木遮蔽(表1),景宁玉兰可获得较充分的光照和空间资源[20],有利于其一级枝生长和提高逐步分枝率,增强其与伴生树种的生存竞争力 。
研究结果表明,景宁玉兰生殖萌枝的花量均与萌枝大小和高度呈正相关关系(图1),但生殖萌枝比例及其花量与每丛最大萌枝的高度没有关系(P>0.05),说明植物开花是以植物体内有机物质的积累为基础,但个体生殖成功与否还受到花期环境条件及传粉昆虫活动状况的综合影响[24,28]。本研究发现,景宁玉兰植株的生殖萌枝花量及其比率与海拔高度有关(表4),随海拔升高而逐渐下降,如从低海拔的杉木林、落叶阔叶林到高海拔的黄山松林,景宁玉兰植株的生殖萌枝花量及其比率呈逐渐下降的趋势;但其结实状况却呈现与开花性状不同的现象,如高海拔黄山松林下景宁玉兰生殖萌枝果实数、单果种子数和每丛平均结实率均明显高于中海拔的落叶阔叶林和杉木林(表3),可能黄山松林下景宁玉兰早春开花时,伴生树种金缕梅同期开花,在吸引传粉昆虫方面发挥了积极作用,说明环境条件是影响景宁玉兰生殖成功与否的重要因素[8,29]。不同植被类型对景宁玉兰的生殖萌枝开花及结实情况也有一定影响(表4),如景宁玉兰在杉木林下的生殖萌枝花量显著高于林缘,林缘生殖枝比例显著高于落叶阔叶林,落叶阔叶林下景宁玉兰没有发现结实现象(表3)。景宁玉兰属先花后叶植物,虽然个体光合有机产物的积累是影响个体生殖构件发育的直接因素,但花果期败育状况与环境条件关系密切[24],有机物质分配策略与雌雄花异熟的生物学特性[29-30]和传粉昆虫多寡及其访花效率均是影响生殖成功的主要因素[31]。
植物体是作为一个整体对外部环境变化做出反应,由于环境变化导致的某个性状变化可能引起其他性状的相应变化,进而影响着植物体对变化环境的整体响应。本研究只是分析比较了不同生境上景宁玉兰萌枝的形态及生殖生物学性状,今后应重点关注微生境变化及传粉昆虫活动对景宁玉兰生态生理及生殖发育方面的影响,探讨影响景宁玉兰生殖成功率的关键因素及其影响机制,深入了解景宁玉兰的生态适应性和种群健康维持机制。
5 结论
(1)不同生境下景宁玉兰萌枝形态特征存在差异。景宁玉兰萌生能力与母株的大小存在一定关系,但萌枝大小和高度与生境更为密切相关。其枝系特征在不同生境上的表现也有较大差异,落叶阔叶林下景宁玉兰萌枝大小及分枝率最低。
(2)不同生境下景宁玉兰萌枝生殖特征存在差异。景宁玉兰生殖萌枝的花量均与萌枝大小及其高度呈正相关关系,生殖萌枝的花量与其结实率没有关系。落叶阔叶林下景宁玉兰生殖枝花量及其结实率均最低,虽然黄山松林下景宁玉兰生殖枝花量较低,但其萌枝结实率、单果种子数及每丛结实率最高。
(3)生境条件对景宁玉兰萌枝形态及其生殖特征存在显著影响。生境类型和海拔高度对景宁玉兰萌枝基径、高度、一级枝长度、生殖萌枝花量、生殖萌枝比率及生殖萌枝结实率均存在显著影响。
致谢:松阳县林业局景宁玉兰小区护林员雷国兴和丽水莲都生态林场江丽荣参加部分野外工作,特此致谢。
猜你喜欢
北京大学学报(自然科学版)(2022年1期)2022-02-21
绿色科技(2021年13期)2021-07-29
花卉(2020年23期)2020-12-08
浙江柑橘(2020年3期)2020-10-12
现代农业科技(2019年17期)2019-10-30
中华魂(2018年11期)2018-05-14
黄山学院学报(2017年5期)2017-11-03
浙江农业科学(2016年11期)2016-05-04
浙江林业(2015年10期)2015-12-11
武夷学院学报(2015年3期)2015-07-18
