
基于叶绿体16S rRNA的绿色裸藻类系统发育及性状进化研究
2019-01-04 05:09:54王艳梅吕俊平南芳茹谢树莲
西北植物学报 2018年11期
王艳梅,冯 佳,吕俊平,刘 琪,南芳茹,谢树莲
(山西大学 生命科学学院,太原 030006)
裸藻类是真核生物演化过程中的一个独立支系,起源较早,绝大多数种类是单细胞,只有极少数是由多个细胞聚集成的不定群体[1-2]。裸藻类植物兼有动植物双重性状,多数种类可以吞噬或胞饮方式吸收营养,具有色素体的种类可进行光合作用。现已有研究表明,绿色裸藻类是通过二次内共生起源的[3-6]。
绿色裸藻的最早发现可以追溯到340多年前,然而,作为一个生物类群进行系统的分类研究却经过了150多年[7]。从19世纪30年代开始,有关绿色裸藻的研究逐渐发展起来,先后报道了裸藻属(Euglena)[8]、隐裸藻属(Cryptoglena)[9]、柄裸藻属(Colacium)[10]、囊裸藻属(Trachelomonas)[10]、扁裸藻属(Phacus)[11]、鳞孔藻属(Lepocinclis)[12]、双鞭藻属(Eutreptia)[13]、旋形藻属(Monomorphina)[14]、拟双鞭藻属(Eutreptiella)[15]和陀螺藻属(Strombomonas)[16],特别是随着分子生物学手段的引入,有关绿色裸藻类分类和系统发育的研究有了进一步深入的发展,相继建立了盘裸藻属(Discoplastis)[17]、眼裸藻属(Euglenaria)[18]和吞噬藻属(Rapaza)[19]。迄今,绿色裸藻类共报道上述13属。
绿色裸藻类有很多共同特征,细胞大多或多或少呈纺锤形,少数为圆柱形、椭圆形或卵形,有的细胞侧扁,无细胞壁,但质膜内有特化的表质,也叫周质体,有的种类胞外具囊壳,细胞前端具有胞口、胞咽、伸缩泡、眼点等结构,具1~2条鞭毛,细胞核较特殊,有丝分裂时,核膜和核仁不消失,色素体形状多样,大小和数目不一,贮藏物质为副淀粉[20]。
长期以来,裸藻类的形态结构是其重要的分类依据[21]。施之新[22]曾基于形态性状对裸藻类的系统发育进行过研究。然而,单纯以形态特征来探究裸藻类植物的系统发育是不完善的,有的种类形态很相似,但遗传距离却很远[18]。以形态特征和分子数据相结合研究绿色裸藻类的系统发育及性状进化很有必要。
叶绿体16S rRNA基因是所有原核和真核生物共同具有的基因,目前GenBank已收录了数万条16S rRNA基因序列,积累了丰富的序列资料。同时16S rRNA 参与细胞的肽链合成,在进化上高度保守,因此是构建进化谱系的理想基因[23]。植物叶绿体中rRNA基因排列顺序与原核生物大肠杆菌和蓝绿藻的rRNA基因排列顺序相似,而且这些rRNA基因大多处于反向重复区内,即有2套方向相反的rRNA基因[24]。由于16S rDNA序列一级结构比较保守,而二级结构又表现出螺旋差异性,较适用于种、属水平的系统学研究[25-26]。
本研究基于叶绿体16S rRNA基因序列,构建了绿色裸藻类的系统发育关系,并对其性状进行了分析和祖先重建,以期探讨绿色裸藻类的系统演化关系,为研究该类植物的起源提供进一步的证据。
1 材料和方法
1.1 分类单位
本研究包含了绿色裸藻类植物12属88种(表1)。
1.2 方 法
1.2.1基因序列在NCBI基因库中搜索绿色裸藻类各属种所包含的全部16S rRNA基因序列,选取各个种对应的一条较长序列用于分析。各序列登录号及对应文献见表1。
1.2.2系统发育分析用BioEdit 7.0.5.3软件进行拼接及校对,以确保准确性,之后运用MrBayes3.2软件构建贝叶斯系统发育树。
1.2.3性状的选择性状选择以选取尽可能多的相对稳定的性状状态和有明显间断性变异的数量性状为原则[27]。性状状态主要根据文献的广泛比较和研究来确定。用于分支分类的性状及状态编码见表2。所有的性状均为无序、等权。代表性状状态的数字没有进化意义。不详或未知的状态用“?”表示(表1)。
网格无关性验证条件为: 喷流Majet=1.0, Tjet=300 K, Pjet=0.96 MPa, 来流Ma∞=2.52, T∞=754 K, P∞=86 kPa. 图2给出了使用3种网格得到的计算模型上壁面压力系数分布. 观察可得: 3种网格所得到的上壁面压力系数变化趋势一致, 其中中等网格和加密网格在数值上基本一致, 即对于本文算例, 中等密度的计算网格即可满足计算要求.
对绿色裸藻类的形态特征进行全面的比较分析后[21,28-35],选取8个性状用于祖先重建。
1.2.4祖先重建分析基于系统发育关系研究中的MP分析,计算10 000 000代得出的10 001棵严格一致树中选取最后的1 000棵树,加入所选取的8个性状(表2),通过Mesquite3.40[36]软件进行形态性状祖先重建。
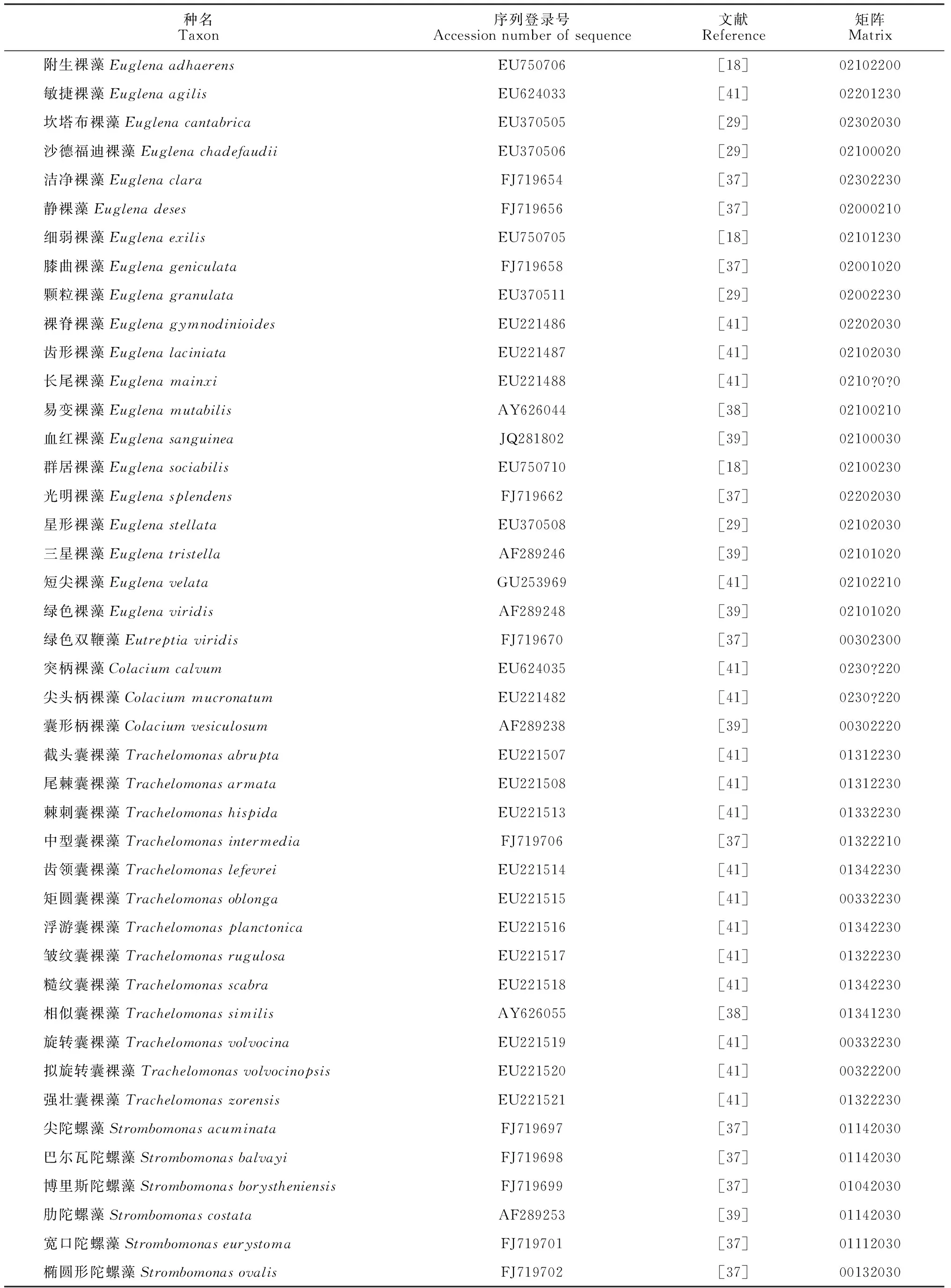
表1 绿色裸藻类序列登录号及形态数据矩阵
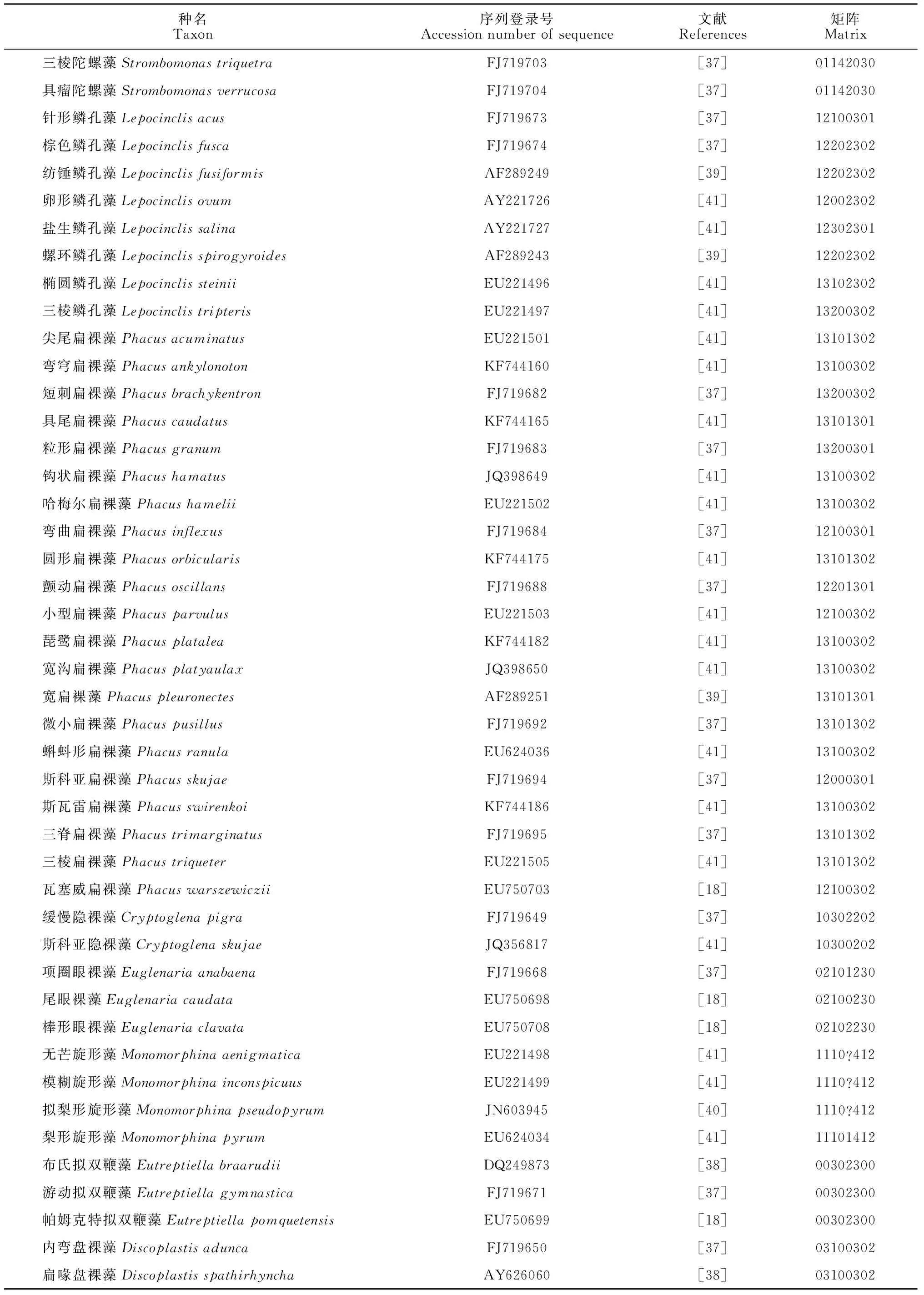
续表1 Continued Table 1

表2 绿色裸藻类形态性状编码表
2 结果与分析
2.1 绿色裸藻类系统发育关系分析
贝叶斯法构建的绿色裸藻类系统发育树(图1)显示,双鞭藻属与拟双鞭藻属互为姐妹群且得到高度支持(100%)。扁裸藻属、鳞孔藻属和盘裸藻属位于一支,也得到了很高的支持率(100%)。囊裸藻属和陀螺藻属也聚为一支,同样也得到了很高的支持率(100%)。裸藻属、隐裸藻属、柄裸藻属和旋形藻属亲缘关系较近。裸藻属不是一个单系类群,在系统发育发面仍需要深入研究。其它个别属虽然也有出现嵌套现象,但整体系统发育关系较清晰。
2.2 绿色裸藻类祖先重建及各形态性状原始性分析
基于形态性状对绿色裸藻类祖先重建的结果(图2~3)显示:(1)表质硬化程度 表质硬化程度在绿色裸藻类不同属表现各异,或是柔软易变形,或是硬化而形状固定。从该性状的祖先状态重建结果(图2)可以看出,表质柔软易变形这一性状较原始。
(2)表质条纹 表质条纹作为绿色裸藻类的常见性状,形态多样,如裸藻属、眼裸藻属、大部分鳞孔藻属和一部分扁裸藻属种类为螺旋形线纹,盘裸藻属和一部分扁裸藻属种类表现为纵线纹,陀螺藻属、旋形藻属和大多数囊裸藻属种类表现为孔纹、点纹、凸纹或肋纹,柄裸藻属、隐裸藻属、双鞭藻属和拟双鞭藻属等少数种类则表面光滑。由该性状的祖先状态重建结果(图2)可以看出,螺旋形线纹是最早出现的原始性状。
(3)后端形状 细胞后端或多或少渐狭是裸藻类典型特征之一,但变化较多。从该性状的祖先状态重建结果(图3)可以看出,细胞后端渐尖或尖尾刺状是最为原始的性状。
(5)鞭毛长度 在绿色裸藻类中,鞭毛长度这一性状特征明显,便于观察和测量,在同一种类中,比较稳定,短于体长,等于体长,或长于体长,可作为典型的分辨性状。由该性状的祖先状态重建结果(图4)可以看出,在发展过程中,3种性状出现频率相近,在可能的祖先状态中各占一定比例,其中鞭毛等于体长这一性状占比较小,鞭毛长于或短于体长2种性状占比相近,原始性不确定。
(6)叶绿体形状 叶绿体形状是绿色裸藻类分类的重要性状,有的为星状,有的较大,为片状、盾状或大盘状,有的较小,为小盘状或球形。由该性状的祖先状态重建结果(图4)可以看出,叶绿体为片状、盾状或大盘状这一性状较原始。

分支节点处数字代表16S rRNA基因贝叶斯系统发育树节点支持率,低于0.50不显示;标尺数字代表遗传距离。图1 基于16S rRNA序列的绿色裸藻类贝叶斯系统发育树The figures of the branch node represent the support rates of Bayesian tree based on 16S rRNA gene, and it is not displayed when lower than 0.50; the number of scale represents genetic distanceFig.1 Bayesian tree of green euglenophytes based on 16S rRNA sequence
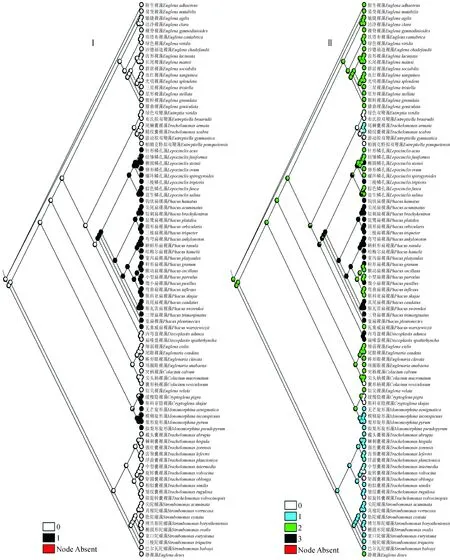
图2 基于表质硬化程度(Ⅰ)和表质条纹(Ⅱ)性状的绿色裸藻类祖先重建结果Fig.2 Results of ancestral reconstruction about green euglenophytes based on degree of surface hardening (Ⅰ) and superficial stripe (Ⅱ)
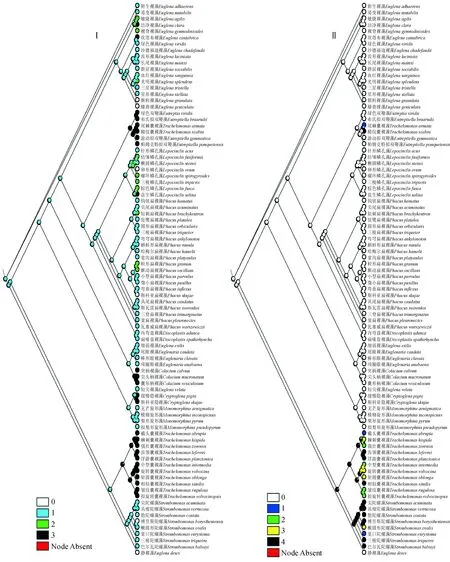
图3 基于后端形状(Ⅰ)和囊壳及鞭毛孔结构性状(Ⅱ)的绿色裸藻类祖先重建结果Fig.3 Results of ancestral reconstruction about green euglenophytes based on shape of posterior end (Ⅰ) and lorica and flagellar pore (Ⅱ)

图4 基于鞭毛长度(Ⅰ)和叶绿体形状(Ⅱ)的绿色裸藻类祖先重建结果Fig.4 Results of ancestral reconstruction about green euglenophytes based on length of flagella (Ⅰ) and shape of chloroplast (Ⅱ)
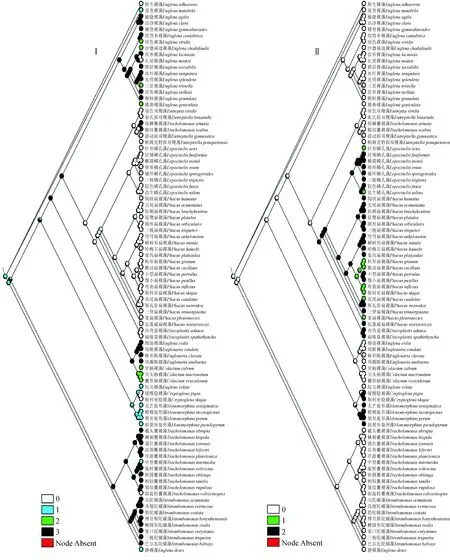
图5 基于叶绿体蛋白核(Ⅰ)和副淀粉粒形状及数量(Ⅱ)性状的绿色裸藻类祖先重建结果Fig.5 Results of ancestral reconstruction about green euglenophytes based on pyrenoid of chloroplast (Ⅰ) and shape and quantity of paramylon grains (Ⅱ)
(7)叶绿体蛋白核 蛋白核是绿色裸藻类色素体中的重要结构,有的种类缺乏,有的种类存在,有蛋白核的种类,有的具副淀粉鞘,有的无。由该性状的祖先状态重建结果(图5)可以看出,无蛋白核这一性状较稳定而经历了长的发展时期,蛋白核具鞘这一性状也在祖先状态中以一定频率出现,但最原始的性状可能是蛋白核无鞘这一性状。
(8)副淀粉粒形状及数量 副淀粉粒形状及数量这一性状表现稳定,是绿色裸藻类种类鉴定的重要特征,表现为小颗粒状且数量不定、具定形但不定数不定位的大颗粒或具定形定数定位的大颗粒。由该性状的祖先状态重建结果(图5)可以看出,副淀粉粒为小颗粒状且数量不定这一性状相对原始。
2.3 基于祖先重建的绿色裸藻类植物原始性分析
李效宇等[42]研究表明,绿色裸藻不同种类的体表线纹在排列方向、形状、粗细及疏密程度不同,而且这一性状在同一定种内是相对稳定的,因此可以作为分类的一个重要依据。本研究通过对表质条纹(图2)的研究发现,对于光滑或不明显,孔纹、点纹、凸纹或肋纹,纵线纹3种性状来说,螺旋形线纹这一性状更为原始,推测表现为螺旋形线纹的裸藻属、眼裸藻属、鳞孔藻属和扁裸藻属有可能是较早出现的绿色裸藻类。通过对表质硬化程度(图2)的研究发现,对于形状固定这一性状来说,易变形的性状更为原始,推测表现为易变形的裸藻属、眼裸藻属、双鞭藻属、拟双鞭藻属、囊裸藻属、陀螺藻属和盘裸藻属有可能是较早出现的绿色裸藻类。对后端形状的研究(图3)发现,对于形状不定、乳头状突出、圆形3种性状来说,渐尖或尖尾刺这一性状更为原始,推测表现为后端渐尖或尖尾刺的裸藻属、眼裸藻属、扁裸藻属、陀螺藻属、盘裸藻属和旋形藻属有可能是较早出现的绿色裸藻类。对囊壳及鞭毛孔的研究(图3)可以看出,对于有囊壳无领,具或无加厚环;无领,具加厚环;具矮领,具或无加厚环;具领,领缘具齿刻四种性状来说,无囊壳这一性状更为原始,推测表现为无囊壳的裸藻属、隐裸藻属、柄裸藻属、扁裸藻属、鳞孔藻属、双鞭藻属、旋形藻属、拟双鞭藻属、眼裸藻属和盘裸藻属出现相对较早,而囊裸藻属和陀螺藻属出现较晚。对鞭毛长度的研究(图4)可知,鞭毛长度在发展过程中,无特定的侧重发展方向,各性状出现频率接近,不能推断可能的祖先状态。通过对叶绿体形状的研究(图4)发现,对于星状、小盘状、球形3种性状来说,片状、盾状或大盘状这一性状更为原始,推测表现为叶绿体片状、盾状或大盘状的囊裸藻属、柄裸藻属、隐裸藻属、眼裸藻属和裸藻属可能较早出现。通过对叶绿体蛋白核的研究(图5)发现,对于无蛋白核、蛋白核被副淀粉颗粒包围、蛋白核具鞘3种性状来说,蛋白核无鞘这一性状更为原始,推测表现为具无鞘蛋白核的裸藻属、囊裸藻属和旋形藻属有可能是比较原始的种类。通过对副淀粉粒形状及数量的研究(图5)发现,对于具定形但不定数不定位的大颗粒、具定形定数定位的大颗粒2种性状来说,副淀粉粒为小颗粒状且数量不定这一性状更为原始,推测表现为副淀粉粒小颗粒状且数量不定的裸藻属、眼裸藻属、双鞭藻属、拟双鞭藻属、囊裸藻属、陀螺藻属和柄裸藻属植物较早出现。综合8种性状祖先重建结果来看,裸藻属和眼裸藻属植物具有所有原始性状,可能是绿色裸藻类的祖先。
3 讨 论
从研究结果看,双鞭藻属与拟双鞭藻属互为姐妹群,鳞孔藻属和盘裸藻属亲缘关系较近,裸藻属不是一个单系类群,这与之前有关文献基于SSU rRNA的研究结果一致[19]。但扁裸藻属、陀螺藻属、隐裸藻属、柄裸藻属和旋形藻属亲缘关系则与文献报道结果有出入。另外,有些不同属亲缘关系很近,同时有的属内不同种类亲缘关系较远,出现这种现象可能是基于不同的基因会出现不同的结果。也有可能目前属种的界定还不够完善。因此,只有在更多分子序列数据的基础上,才可能构建科学客观的系统发育关系。
不同物种性状的形态稳定性对于重建复杂性状的进化史是必要的,以形态性状和分子数据相结合重建性状的发展过程,是科学发展的必然[19]。施之新曾基于形态性状,对裸藻类进行过分支分类分析,指出对于表质的硬化程度及变形这一性状的极性是沿柔软变形向高度硬化而不变形的方向发展,对于表质条纹这一性状的极性是由螺旋形向纵向过渡,对于囊壳及鞭毛孔这一性状的极性是由无囊壳向囊壳具领和加厚环过渡,对于副淀粉粒这一性状的极性是由小颗粒状且数量不定向具定形定数定位的大颗粒过渡[22]。本研究结果与其高度一致,可见基于叶绿体16S rRNA可以客观反映绿色裸藻类的进化过程。
本研究所选择的叶绿体16S rRNA基本上可以客观反映绿色裸藻类系统发育关系,说明该基因是植物类群系统发育的有效分子标记之一,也说明了叶绿体基因在反映植物系统发育研究中的重要性。但是采用单一的分子标记也存在一定的缺陷。由于分子数据的局限以及分析方法存在一定程度的不足,因此祖先重建分析的结果可能只是基本上反映祖先性状的演化过程。此外,还有一些其它性状如细胞形状、囊壳的分层以及囊壳的颜色等,虽然本研究尚未涉及,但是这些性状在绿色裸藻类植物分类方面依然具有一定的价值。因此,进一步积累分子数据,更为合理地对形态性状进行编码,进一步正确评价性状及其分类学或系统学意义,值得进一步深入研究。
猜你喜欢
法医学杂志(2022年1期)2022-06-21 01:23:34
科学大众(2021年21期)2022-01-18 05:53:38
小哥白尼(趣味科学)(2018年10期)2019-01-16 01:32:20
红领巾·探索(2018年10期)2018-11-14 02:49:28
中国环境科学(2018年7期)2018-07-26 09:03:50
法医学杂志(2016年5期)2016-11-21 01:44:43
广西林业科学(2016年1期)2016-03-20 05:33:01
中国环境科学(2015年6期)2015-11-19 08:39:58
课堂内外(初中版)(2015年12期)2015-09-10 07:22:44
食品科学(2013年6期)2013-03-11 18:20:13
