
北京松山自然保护区典型植物群落物种多样性研究
2019-01-04 05:10鲁绍伟李少宁吴记贵范雅倩
西北植物学报 2018年11期
赵 娜,鲁绍伟*,李少宁,吴记贵,范雅倩
(1 北京市林业果树科学研究院,北京 100093;2 北京燕山森林生态系统定位观测研究站,北京 100093;3 北京松山国家级自然保护区管理处,北京 102115)
植物群落的物种多样性既反映了环境与植物群落之间的相互关系[1],也体现了各物种在群落中的优势度、均匀度和丰富度[2],对其研究可深入了解群落物种的组成、群落变化及发展趋势,乃至预测生态系统的功能发挥[3],因此植物群落的物种多样性及其与生态环境、群落和生态系统稳定性的相互关系已成为生态学的研究重点[4]。α与β多样性共同构成了群落总体多样性或者一定地段的生物异质性。而α与β多样性指数并不一定发生协同变化[5]。β多样性为沿环境梯度变化物种替代的程度[6],同时也可反映群落物种组成的差异。不同β多样性指数具有不同的指示意义。二元属性测度的β多样性指数包括Whittaker指数(βws)和Cody指数(βC)。βws可直观反映β多样性与物种丰富度之间的关系。βC则体现沿生境变化增加或失去物种数目。二元属性β多样性测度计算简单且易于使用,但未考虑群落中每个个体的相对多度,导致对稀有种的作用过高评价。而数量数据β多样性测度,能够弥补二元属性β多样性测度的不足,以分析不同群落间的相似性系数。本研究采用Jaccard指数(CJ)、Sorenson指数(CS)和Bray-Curtis指数(CN)3种数量数据β多样性指数共同分析群落间的β多样性差异。
设立自然保护区仍然是保护生物多样性的核心举措[7],是实现生物多样性及其生态系统服务的重要途径。北京松山国家级自然保护区,属华北地区保存比较完好的代表性暖温带山地森林生态系统,为北京地区防沙治沙起着至关重要的作用。松山自然保护区物种丰富、群落结构复杂[8]。1986年建区初期,由于缺乏重视与管理落后,资源过度开发,生物多样性受到严重威胁。目前已有针对松山自然保护区的人为干扰、生态旅游、环境教育与保护、森林资源及其服务价值评估等方面开展相关研究[9-11]。地形、海拔等所引发环境综合作用共同控制物种组成与多样性的空间分布特征[12-14]。因此,不仅要注重群落物种组成和多样性保护,同时探讨变化环境造成群落间多样性差异也至关重要。
因此,本研究在北京松山自然保护区、海拔高度700~1 000 m处,根据典型植被类型设置6块标准样地,调查植物群落乔木、灌木和草本层物种组成与α、β多样性特征,为进一步分析森林生态系统功能发挥以及自然保护区后续建设与保护措施的制定提供一定的数据支撑。
1 材料和方法
1.1 研究区概况
研究固定样地设置于北京市延庆区西北部的松山国家级自然保护区界内,距市区90 km(E115°43′44″~115°50′22″,N40°29′9″~40°33′35″)。该自然保护区西、北部分别与河北省赤城县大海坨国家级自然保护区相接,其西南部与河北省怀来县接壤,东部与北京市玉渡山自然保护区毗邻,南部与延庆区张山营镇相邻。松山国家级自然保护区总面积4 671 hm2,森林覆盖率87.65%。属暖温带大陆性季风气候区。年均温6~8.5 ℃,年均日照时数2 500 h,≥10 ℃积温为2 500 ℃左右,无霜期140 d,年降水量470 mm,年蒸发量约1 600 mm。
1.2 样地设置
2017年7月,遵循样方布设与周围环境条件一致、避开林缘地带的原则,采用典型样方法,于北京松山自然保护区海拔700~1 000 m段,在其6个典型林分类型区内分别设置1块标准样地(60 m×60 m),在每个标准样地内分别设3块20 m×20 m乔木样方;在每个乔木样方内,随机设置2块5 m×5 m的灌木样方、3块1 m×1 m的草本样方。所选样地区域内包含该保护区内典型地带性乔木油松(Pinustabulaeformis)、蒙古栎(Quercusmongolica)、山杨(Populusdavidiana)和毛白杨(Populustomentosa)等(表1)。
1.3 调查方法
对乔木样方内所有乔木层植被予以编号,并记录乔木层植被种名、株数、胸径(1.3 m胸径)、株高、冠幅(四向冠幅均值)及枝下高等。每个灌木、草本样方内,测定并记录灌木层、草本层物种种名、数量、高度、冠幅、盖度和频度[15]。

表1 北京松山自然保护区标准样地基本概况
1.4 数据分析
1.4.1重要值测度计算固定样地内乔、灌、草物种的重要值:
重要值=(相对频度+相对盖度+相对密度)/3
(1)
1.4.2多样性测度本研究分别采用α和β多样性测度分析松山植物物种多样性的空间分布特征[5]。
①α多样性指数
本研究选取的α多样性指数包括Simpson指数(H′)、Shannon-Wienner指数(H)、Margalef丰富度指数(R)、Pielou均匀度指数(Js和Jsw)和Alatalo均匀度指数(Eu),计算公式如下:
(2)
Shannon-Wienner多样性指数:
(3)
Margalef丰富度指数:
R=(S-1)/lnPi
(4)
Pielou均匀度指数(包含Js和Jsw):
(5)
Jsw=H/lnS
(6)
Alatalo均匀度指数:
(7)
式中,Pi=Ni/N表明第i种的相对重要值;Ni是第i种的重要值;N为全部物种的个数之和;S表示样方中的物种总数。
②β多样性指数
本文选用的β多样性指数,包括基于二元属性数据的Cody指数(βc)、基于数量数据的相似性系数Jaccard和Sorenson指数(CJ和CS指数)、Whittaker指数(βws)以及Bray-Curtis指数(CN),计算公式如下:
Cody指数:βC=(b+c)/2
(8)
Jaccard指数:Cj=a/(a+b+c)
(9)
Sorenson指数:CS=2a/(2a+b+c)
(10)
式中,a为2个研究样地中的共有物种数目;b是沿生境梯度所失去的物种数目,即上一个梯度内存在但在下一个梯度没有的物种数目;c指沿生境梯度所增加的物种数目,即上一个梯度不存在而在下一个梯度内存在的物种数目。
Whittaker指数:βWS=S/ma-1
(11)
式中,S为研究样地内所记录的物种总数;ma是各样方中平均物种数。
Bray-Curtis指数:CN=2jN/(Na+Nb)
(12)
式中,Na为样地A内各物种所有个体数目和;Nb是样地B中物种所有个体数目和;jN指样地A和B中共有种个体数目较小者之和:
jN=∑min(jNa,jNb)
(13)
1.4.3统计分析利用SPSS18.0进行单因子方差分析。对各因子进行Pearson相关性分析。采用Excel 2010对试验数据进行整理、作图分析。
2 结果与分析
2.1 物种组成及数量特征
据调查统计,6个植被类型样地内共有75种植物种,隶属40科58属。其中,菊科(Compositae)最多,蔷薇科(Rosaceae)和堇菜科(Violaceae)其次,榆科(Ulmaceae)、茄科(Solanaceae)、壳斗科(Fagaceae)和茜草科(Rubiaceae)物种数目较少(仅1种),但其多度较大(>20)(表2)。乔木树种隶属10科11属11种,除3号样地外,其他样地乔木优势种显著。3号样地为杂木林,其中乔木优势种为毛白杨(占样地乔木种总数的30%),次优种为胡桃楸(25%)。灌木物种较少,共11科12属13种(表2和3),重要值较大的物种有土庄绣线菊(Spiraeapubescens)、扁担杆(Grewiabiloba)、荆条(Vitexnegundovar.heterophylla)和大花溲疏(Deutziagrandiflora)。由本研究区重要值前5名草本物种名录(表4)可知,草本物种24科40属51种,多中生或阳生植物,以菊科草本分布最广,种类最多。除3号样地阴生植物居多外,各样地草本均以阳生和中性植物为主。主要草本物种有三脉紫菀(Asterageratoides)、大头橐吾(Ligulariajaponica)、野海茄(Solanumjaponense)、透骨草(Phrymaleptostachyasubsp.asiatica)、黄瓜假还阳参(Crepidiastrumdenticulatum)、异鳞苔草(Carexheterolepis)、大丁草(Gerberaanandria)和叉岐繁缕(Stellariadichotoma)等(重要值大于15%)。
2.2 物种多样性和植物生长型的关系
不同植物生长型H′、H和R变化趋势为草本>灌木>乔木,其中3号杂木林样地乔木、灌木的H′、H和R均显著高于其他样地,但其草本R却低于其他样地(图1);且1、2和3号样地灌木H′、H和R均高于4、5和6号样地。由图1可知,1、2和4号样地内乔木Eu相对较低(0.44、0.41和0.40),且4号样地灌木各均匀度指数(0.48)远低于其他样地。各个样地草本均匀度间差异较小。综上,除3号样地乔木组成丰富外,本研究区域其他样地乔木物种组成单一。随着海拔高度增加,灌木多样性与丰富度指数减小,草本丰富度指数略增,乔、灌和草均匀度随山体高度增加无显著变化。

表3 北京松山自然保护区6种植被类型样地主要灌木物种重要值
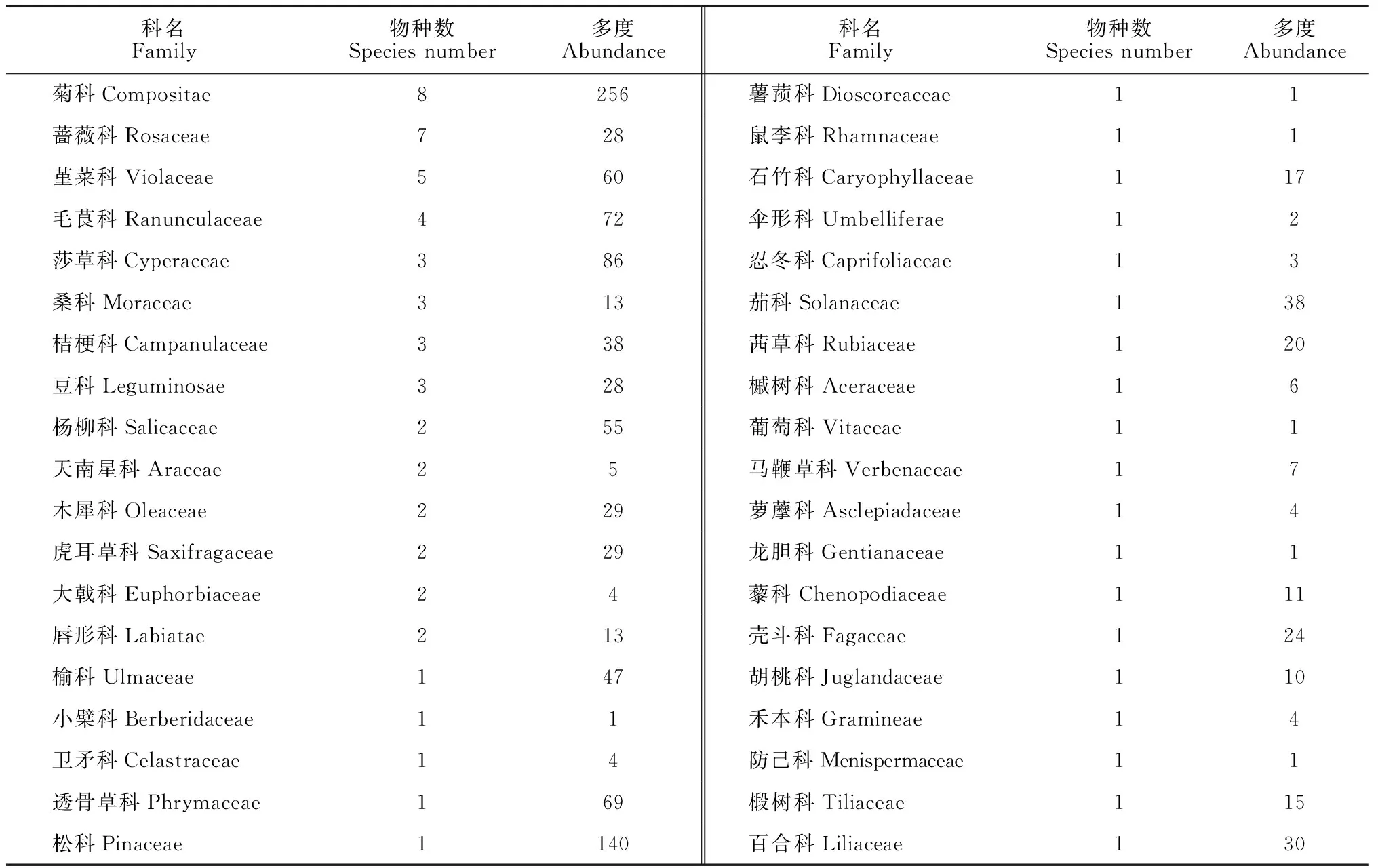
表2 北京松山自然保护区6种植被类型样地各植物种组成分析

表4 北京松山自然保护区6种植被类型样地重要值前5位草本物种
注:大叶白蜡、桑和榆处于幼苗期时,将其归于草本样方统计,其幼树统计于灌木样方内,其成年乔木归于乔木样方统计。幼年南蛇藤和花木蓝属草本样方调查范围,其成年个体统计于灌木样方内
Note:F.rhynchophylla,MorusalbaandU.pumilaat seedling stage were surveyed in the plots of herb layer, whose young trees were calculated for shrub layer and their adult trees belonging to tree layer.CelastrusorbiculatusandIndigoferakirilowiiat young period were observed for herb layer, which at mature period were counted for shrub layer

图1 北京松山自然保护区不同生长型植物多样性Fig.1 Plant diversity between different growth types in Songshan Nature Reserve, Beijing
2.3 林下草本物种多样性
2.3.1草本物种α多样性比较保护区林下草本α多样性发现,随着山体高度的增加,除2号样地油松林外,其他样地林下草本Shannon-Wienner多样性指数(H)逐渐递增,而Simpson多样性指数(H′)无显著差异(图2);4号山杨林样地草本物种丰富度指数最高(4.41),3号样地杂木林最低(2.50)。随着山体高度增加,各Eu、JS与JSW指数均增大。1号蒙古栎林下草本均匀度指数最高,而2号样地则最低,这可能与油松林下草本物种种类稀少相关。
2.3.2草本物种β多样性从草本群落相异性角度分析6块样地林下草本β多样性可知,1~6样地间βC指数最大(15.50),且随着山体高度的增加,相邻样地间草本βC指数呈先增加后减小趋势(表5)。3号样地与其他样地间的草本βWS较大,即草本共有种较少,且相邻样地间草本βWS与βC指数随海拔变化趋势相似;其中2~3样地间βWS指数最大(3.14),4~6样地间最小(1.83)。

图2 北京松山自然保护区林下草本物种α多样性比较Fig.2 Alpha diversity of understory herbaceous species in Songshan Nature Reserve, Beijing
由表5可知,CJ和CS反映各样地草本β多样性变化趋势基本一致。3号杂木林草本物种与其他样地间的相似度较低,物种间隔度大,表现为2~3号样地间CJ、CS和CN最低。相邻两样地间CJ、CS和CN均表现为随着海拔高度递增。2号、5号和6号油松林下草本CJ、CS和CN较大,共有种较多,可见草本相似性β多样性受上层乔木组成的影响。
3 讨 论
3.1 物种组成及数量特征
除3号杂木林样地外,其他样地乔木组成相对单一;灌木多以土庄绣线菊和扁担杆为优势种;本地区林下草本植物丰富度与多样性均较高。调查表明随着山体高度的增加,三脉紫菀的重要值逐渐减小,这说明海拔作为一个复合环境因子,会影响上层乔木叶面积指数,二者协同影响林下草本组成结构,这与朱源等[16]研究贺兰山林下草本物种组成随海拔的变化趋势一致。
杂木林的多乔木特征与草本物种多样性往往呈负相关,即为抑制作用[17]。3号样地杂木林中的乔木层和灌木层物种种类最丰富,但草本层物种种类较其他样地少,且多以阴生植物为主,这正与林冠较大的叶面积指数相对应(3.51),即林冠郁闭可导致喜阴草本植物占优势地位。而其他各样地林下均分布有菊科草本,且重要值较高,这与赵勃[18]所研究的北京山区植被分布的结果一致。由此进一步证实菊科草本生态位广,资源利用能力强,在北京山区分布广泛,能够较好地适应松山海拔700~1 000 m环境条件;同时也说明物种重要值与生态位具有正相关性[19-20]。
3.2 物种多样性和植物生长型的关系
林下环境复杂,则林下草本物种多样性特征表现有所不同[21],由此造成本研究区草本层物种丰富度和多样性均大于乔、灌木层,这与张建宇等[17]的研究结果一致。对比各样地不同植物生长型灌木多样性、丰富度指数发现,1、2和3号样地的灌木物种多样性、丰富度与均匀度指数均大于其他样地,但其草本层植被多样性却不及后3个样地,可能与空间异质及演替过程对生境的影响作用相关[22],也存在灌木多样性大于草本的现象[19]。与此同时,还需要综合考虑不同林型所带来林下植被差异[17]。
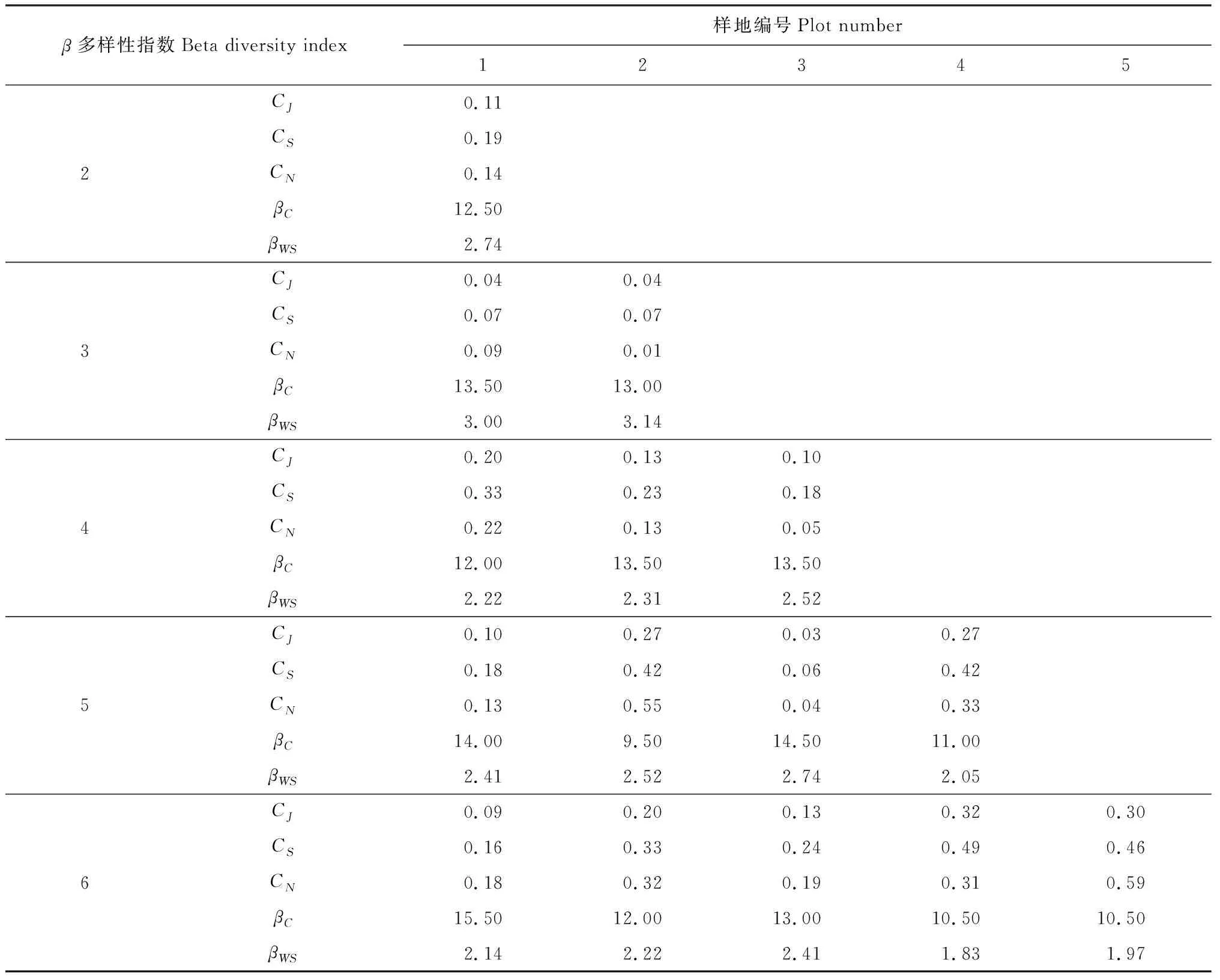
表5 北京松山自然保护区林下草本物种β多样性指数
3.3 林下草本物种多样性特征
陈煜等[23]研究发现,温带森林林下草本物种占群落全部维管植物的90%以上。这就说明林下草本物种不仅可指示立地环境,同时其还具有强大的生态功能予以发挥[24]。在分析林下草本物种多样特征时,要综合考虑冠层物种组成与结构、微地形以及土壤因子的调控作用[25]。综合分析本研究区林下草本多样性、丰富度与均匀度指数发现,随着山体高度的缓慢上升,林下草本多样性与均匀度指数呈增大趋势,这是由于本研究区位于海拔717~950 m范围内,正处于丰富度与多样性指数递增区间内[26-27],这与“中间膨胀”规律并不矛盾[28]。而2号和3号样地较大的灌木多样性会对阳生草本产生遮蔽,不利于其生长[29],使草本多样性指数略小。
3.4 林下草本层β多样性变化特征
探讨相邻样地间的各草本β多样性指数发现,相邻两样地间草本物种CJ、CS和CN在2~3号样地间达到最低,后随着海拔逐渐增加(表5)。松山早期砍伐原有植被,次生林随山体自然演替,造成中海拔高度立地条件逐渐复杂化,形成杂木林(3号样地),共有种减小,物种替代率增大,因此该样地林下草本物种与其他样地间的相似度较低,数量数据β多样性指数较低;而二元属性β多样性βWS与βC均在2~3、3~4样地最大,从山底到山顶βWS与βC经历先增加后减小,这与徐广平等研究的从阴坡-滩地-阳坡草本β多样性的变化趋势相一致[30]。尽管2号与3号样地毗邻,但由于乔木组分不同,导致林下环境因子存在差异,共有种缺乏,草本丰富度较低,因此2~3号样地间草本βWS指数中最高。以上现象说明在土壤、光照等立地条件适中条件下,具有较大的环境异质性,因此会出现沿着山体高度增加,林下草本β多样性表现为先减小后增大的变化趋势,这与Gracia等[31]的研究结果相似。
4 结 论
由于其特殊的地理位置,北京松山国家级自然保护区在首都生态环境保护中发挥重要作用,但由于前期缺乏重视与管理落后,造成大量乔木砍伐,资源过度开发,生物多样性受到严重威胁。经过近年来保护力度的加强,砍伐迹地萌生的次生林逐渐成林。结合本研究植物物种多样性特征分析,在今后生物多样性保护实施过程中,需按照自然演替规律,综合考虑冠层物种组成与结构、微地形的调控作用,寻找适合林分类型的森林群落多样性,制定合理可行的保护经验和技术,为森林群落生态功能的发挥提供科学支撑。
猜你喜欢
林业勘查设计(2022年1期)2022-02-15
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
青年文学家(2021年16期)2021-08-05
少儿美术(2020年8期)2020-11-06
现代园艺(2017年13期)2018-01-19
中学理科园地(2016年6期)2017-01-03
小说月刊(2014年1期)2014-04-23
