
7种秋海棠叶片斑纹结构及遗传特性分析
2019-01-04 02:08杜文文马璐琳瞿素萍贾文杰王祥宁崔光芬
西北植物学报 2018年11期
杜文文,段 青,马璐琳,瞿素萍,贾文杰,王祥宁,崔光芬
(云南省农业科学院花卉研究所/国家观赏园艺工程技术研究中心, 昆明 650205)
秋海棠(Begoniaspp.)是秋海棠科秋海棠属多年生草本植物,喜温暖潮湿环境,不耐寒,惧阳光直射,主要分布在非洲、中南美洲和亚洲的热带和亚热带地区,全世界约有1 900多个种[1],15000多个园艺品种[2]。按其观赏性可分为观花类、观叶类和花叶兼赏类。秋海棠植株形态、颜色丰富多样,尤其在叶色方面较为突出,叶片常见白、深绿、黄绿、银、铜红、深紫红及各种复色。复色叶片有2种以上颜色,多呈不规则分布,形成特殊的图案或花纹,其丰富多彩的叶色在观赏植物中并不多见[3]。目前对秋海棠的研究主要集中在资源调查与评价[4-6]、形态解剖学[7-8]、细胞学[9-10]、系统进化[11]等方面,针对秋海棠叶片斑纹的研究分析则相对较少,Zhang等[12]和Sheue等[13]通过表皮光学特性、叶片组织结构观察,叶绿素含量及Fv/Fm测定研究了具金属光泽叶斑的大王秋海棠、白色点状叶斑的水鸭脚秋海棠和掌叶秋海棠、白色不规则片状叶斑的变色秋海棠、绿色块状叶斑的B.chlorostictaSands等 9种秋海棠叶片斑纹的形成。本试验选择叶片具斑纹的7个秋海棠野生种或商业品种为材料,通过观察叶片组织结构,测定叶绿素含量及叶绿素荧光参数分析其叶斑形成原因,并通过杂交后代的表型分离规律初步探索白色斑点的遗传特性,旨在为秋海棠叶片斑纹的深入研究奠定一定的理论基础。
1 材料和方法
1.1 材 料
4种野生秋海棠包括银点秋海棠(B.deliciosaLinden ex Fotsch)、铺地秋海棠(B.handeliivar.prostrata(Irmsch.)Tebbitt)、假厚叶秋海棠(B.pseudodryadisC. Y. Wu)和彩纹秋海棠(B.masonianavar.maculataS. K. Chen, R. X. Zheng & D.Y. Xia),3个秋海棠属商业品种为‘皮卡’(‘Oeympica’)、‘虎斑’(‘Tiger’)及‘非洲丛林’(‘African Jungle’)。研究材料均种植于云南省农业科学院花卉研究所球宿根花卉育种基地。每份材料均为扦插繁殖而成,个体自幼叶即携带叶斑,且随着叶片生长叶斑颜色加深,面积增大,至叶片成熟时叶斑形状和颜色稳定。其中,银点秋海棠、铺地秋海棠和‘非洲丛林’叶片具白斑,假厚叶秋海棠和‘皮卡’叶片为绿斑,彩纹秋海棠和‘虎斑’斑区颜色为褐色。叶片结构观察、叶绿素含量及叶绿素荧光参数测定均取自成熟叶片斑区和非斑区。
1.2 方 法
1.2.1组织结构观察半薄切片的制作参照巩玥等[14]的方法, 样品先后经过2.5%戊二醛和1%锇酸的固定后依次在30%、50%、70%、80%、90%、95%、100%乙醇中脱水各1次,100%乙醇二次脱水,再经过100%丙酮脱水进行包埋,包埋聚合后可切片。切片厚度为1 μm,甲苯胺蓝染色,显微镜观察并照相。
1.2.2超微结构观察方法同半薄切片,切片厚度50~70 nm,醋酸双氧铀先染5~10 min,冲洗,柠檬酸铅后染5 min,再冲洗,烘烤灯烘烤后透射电镜观察、拍照。
1.2.3叶绿素含量测定叶绿素含量测定参照李合生等[15]的方法,分别称取银点秋海棠、铺地秋海棠、假厚叶秋海棠、‘皮卡’和‘非洲丛林’斑区和非斑区叶片,将取好的叶片置于研钵中,加入少量石英砂和碳酸钙粉以及95%乙醇溶液,研磨至匀浆;经过滤、冲洗后装入容量瓶用乙醇溶液定容;以分光光度计测量叶绿素溶液在665、649和470 nm处的吸光度,计算叶绿素浓度。每个样本重复6次,浓度值取平均值。
1.2.4叶绿素荧光参数Fv/Fm值测定运用叶绿素荧光成像仪测定叶片斑区和非斑区的荧光参数。取各种类成熟带斑叶片,锡箔纸包裹叶片暗处理20 min,测定暗适应状态下的最小荧光(Fo)、暗适应状态的最大荧光(Fm),Fv=Fm-Fo,重复9次。
1.2.5叶斑遗传性以银点秋海棠与水鸭脚秋海棠(B.formosana(Hayata) Masam.)、长翅秋海棠(B.longialataK. Y. Guan & D. K. Tian)、光滑秋海棠(B.psilophyllaIrmsch.)进行杂交,杂交组合为:银点秋海棠(♀)×水鸭脚秋海棠(♂),银点秋海棠(♀)×长翅秋海棠(♂),光滑秋海棠(♀)×银点秋海棠(♂),以及银点秋海棠自交。杂交、自交均于2016年9月进行,12月收集杂交种子,2017年3月进行播种,出苗后即观察后代植株的斑纹稳定性,2018年5月统计后代植株斑纹携带株数,每个组合均统计由3个果实播种获得的后代群体,计算带斑和不带斑植株的平均数。
2 结果与分析
2.1 叶片表型、组织结构与超微结构
表型观察发现银点秋海棠(图1,A)、铺地秋海棠(图1,B)和‘非洲丛林’(图1,C)为白色点状叶斑,假厚叶秋海棠具淡绿色块状和条状叶斑(图1,D),‘皮卡’有银绿色环状叶斑(图1,E),彩纹秋海棠为褐色马蹄状叶斑(图1,F),‘虎斑’叶色主要为红褐色(图1,G),并将绿色分割成不规则块状,本文暂定‘虎斑’斑区颜色为红褐色,另外,银点秋海棠和铺地秋海棠每个斑点中央有一个毛状体,而其他种类则没有。
半薄切片观察到7种秋海棠叶片均由5层细胞组成,分别为上下表皮细胞各1层,栅栏组织细胞1层和海绵组织细胞2层。各种类斑区和非斑区上表皮细胞大小不一,但形状基本为扁平或纵向长方形,并偶有略微突起;下表皮细胞形态与上表皮细胞类似,其中‘虎斑’斑区和非斑区上表皮细胞上方还有1层突起。银点秋海棠、假厚叶秋海棠、铺地秋海棠、‘非洲丛林’和 ‘皮卡’斑区栅栏组织细胞呈不规则漏斗状或圆形,排列疏松,内含叶绿体,与上表皮细胞之间存在空隙,海绵组织细胞呈不规则圆形,含有少量或极少量叶绿体;非斑区栅栏组织细胞则呈漏斗状或圆柱状,比斑区略长,排列紧密,内含丰富叶绿体,与上表皮细胞之间无空隙,海绵组织细胞与斑区类似,叶绿体含量较非斑区多(图2,A-1、A-2、B-1、B-2、C-1、C-2、D-1、D-2、E-1、E-2)。彩纹秋海棠和‘虎斑’斑区和非斑区栅栏组织细胞呈圆柱状,与其他5种秋海棠非斑区的栅栏组织细胞相似,含有大量的叶绿体,斑区和非斑区上表皮细胞和栅栏组织细胞间均紧密相连,不存在空隙,彩纹秋海棠斑区和非斑区海绵组织细胞排列疏松,含少量叶绿体(图2,F-1、F-2),‘虎斑’斑区海绵组织细胞排列疏松,非斑区则排列较紧密,亦含有少量叶绿体(图2,G-1、G-2)。

A. 银点秋海棠;B. 铺地秋海棠;C.‘非洲丛林’;D.假厚叶秋海棠;E.‘皮卡’;F. 彩纹秋海棠;G.‘虎斑’ 图1 7种具叶斑秋海棠A. B. deliciosa; B. B. handelii var. prostrata; C. ‘African Jungle’; D. B. pseudodryadis; E. ‘Oeympica’; F. B. masoniana var. maculata; G. ‘Tiger’Fig.1 Seven Begonia species with foliar variegation
通过超薄切片可观察到7种秋海棠斑区和非斑区叶绿体超微结构形状呈不规则椭圆形或细长形,结构均完整,内有丰富的类囊体,基质和基粒片层清晰,银点秋海棠斑区和非斑区基粒片层均较厚(图3,A-1、A-2),假厚叶秋海棠斑区基质和基粒片层较非斑区略薄(图3,B-1、B-2);同时,各种类均含有淀粉粒和脂肪球,尤其是铺地秋海棠斑区和非斑区含量大量的脂肪球(图3,C-1、C-2),彩纹秋海棠斑区和非斑区则有较多的淀粉粒(图3,D-1、D-2)。

A. 银点秋海棠; B. 假厚叶秋海棠;C. 铺地秋海棠;D.‘皮卡’;E.‘非洲丛林’;F.‘虎斑’;G. 彩纹秋海棠;1. 斑区;2. 非斑区;Ead. 近轴表皮细胞;Eab. 远轴表皮细胞;P. 栅栏组织细胞;Sp. 海绵组织细胞;C. 叶绿体;IS. 空隙;Bu. 突起图2 银点秋海棠、假厚叶秋海棠、铺地秋海棠、‘皮卡’、‘非洲丛林’、‘虎斑’及彩纹秋海棠斑区和非斑区组织结构A. B. deliciosa; B. B. pseudodryadis; C. B. handelii var. prostrata; D. ‘Oeympica’; E. ‘African Jungle’; F. ‘Tiger’; G. B. masoniana var. maculata; 1. Variegated areas; 2. Non-variegated areas; Ead. Adaxial epidermis; Eab. Abaxial epidermis; P. Palisade tissue cell; Sp. Spongy tissue cell; C. Chloroplast; IS: Intercellular space; Bu: BumpleFig.2 Leaf structure of variegated areas and non-variegated areas of B. deliciosa, B. pseudodryadis, B. handelii var. prostrata, ‘Oeympica’, ‘African Jungle’, ‘Tiger’ and B. masoniana var. maculata

A. 银点秋海棠;B. 假厚叶秋海棠;C. 铺地秋海棠;D. 彩纹秋海棠;1. 斑区;2. 非斑区;C. 叶绿体;S. 淀粉粒;Sl. 基质片层;Gl. 基粒片层;Lb. 脂肪球图3 银点秋海棠、假厚叶秋海棠、铺地秋海棠及彩纹秋海棠斑区和非斑区超微结构A. B. deliciosa; B. B. pseudodryadis; C. B. handelii var. prostrata; D. B. masoniana var. maculata; 1. Variegated areas; 2. Non-variegated areas; C. Chloroplast; S. Starch; Sl. Stroma lamella; Gl. Grana lamella; Lb. Lipid ballFig.3 Chloroplast structure of variegated areas and non-variegated areas of B. deliciosa, B. pseudodryadis, B. handelii var. prostrata and B. masoniana var. maculata
2.2 叶绿素含量及荧光参数Fv/Fm值
表1显示,银点秋海棠、假厚叶秋海棠、‘皮卡’和‘非洲丛林’斑区的叶绿素a、b及总叶绿素含量均低于非斑区,其中叶绿素总含量比非斑区分别低6.63%、32.5%、29.52%、5.24%,而铺地秋海棠斑区和非斑区差别不大。其中,假厚叶秋海棠斑区叶绿素a及总叶绿素含量与非斑区存在显著差异,‘皮卡’斑区叶绿素a、b及总叶绿素含量与非斑区均存在显著差异。假厚叶秋海棠、‘皮卡’斑区栅栏组织细胞和海绵组织细胞中叶绿体的密度、含量均明显低于非斑区。5种秋海棠中,银点秋海棠斑区和非斑区的叶绿素含量均为最高值,斑区为1.169 mg·g-1,非斑区为1.252 mg·g-1。斑区叶绿素含量最低的种类为‘非洲丛林’,仅0.651 mg·g-1,非斑区则以假厚叶秋海棠的叶绿素含量(0.486 mg·g-1)最低(表1)。虽然假厚叶秋海棠斑区和非斑区颜色较深,但其叶绿素含量均不是最高值,此结果可能与其红色叶背有关。
7种秋海棠离体条件下斑区和非斑区的叶绿素荧光参数Fv/Fm值见表1。除假厚叶秋海棠的斑区Fv/Fm值小于非斑区外,其余6种秋海棠均为斑区高于非斑区。银点秋海棠、铺地秋海棠和‘非洲丛林’Fv/Fm值在斑区与非斑区之间存在显著差异,‘皮卡’斑区和非斑区无显著差异,假厚叶秋海棠斑区Fv/Fm值略低于非斑区,但差异不显著。彩纹秋海棠和‘虎斑’斑区均为红色,非斑区为绿色,彩纹秋海棠斑区Fv/Fm值和非斑区差别不大,而‘虎斑’斑区Fv/Fm值显著高于非斑区,并且其斑区Fv/Fm值为7个种类中的最高值。银点秋海棠、‘皮卡’、‘非洲丛林’和假厚叶秋海棠斑区叶绿素含量低于非斑区,但其叶绿素荧光参数Fv/Fm值高于非斑区或与非斑区无显著差异,说明非绿色的斑区可能同样具有较高的光能利用率。
2.3 叶斑遗传特性
调查结果表明,银点秋海棠杂交、自交组合的后代均发生性状分离,产生有斑植株和无斑植株,其中自交平均每个果实获得有斑植株287株,无斑植株100株,有斑和无斑植株比例近3∶1。银点秋海棠和水鸭脚秋海棠杂交后代中平均每个果实获得有斑植株176株,无斑植株162株,有斑和无斑植株约为1∶1,银点秋海棠和长翅秋海棠杂交则获得有斑植株156株,无斑植株128株,光滑秋海棠与银点秋海棠杂交获得杂交个体在4个组合中最少,有斑植株110株,无斑植株95株,后两个组合平均每个果实获得的有斑植株和无斑植株也接近1∶1(表2)。4个组合播种同一苗床,栽培管理一致,排除外界环境因素,由于银点秋海棠和水鸭脚秋海棠原产台湾,而长翅秋海棠和光滑秋海棠原产大陆,且4个种的基因组大小为水鸭脚秋海棠(2.554 pg)>银点秋海棠(2.180 pg)>长翅秋海棠(0.956 pg)>光滑秋海棠(0.637 pg),所以杂交亲和程度可能与原产地和基因组大小有关。另外,自交、杂交后代中有斑植株除斑纹大小、形状与银点秋海棠类似外,其他的性状尤其在叶形上与银点秋海棠更相似,而无斑植株的整体性状则与无斑亲本更加相像。由于4个组合中,3个杂交后代有斑和无斑植株比较均接近1∶1,自交后代有斑和无斑植株比例约3∶1,根据孟德尔遗传定律,推测银点秋海棠叶片白色点状斑与无斑是1对可遗传的相对性状,白色点状斑是显性性状,无斑是隐性性状。
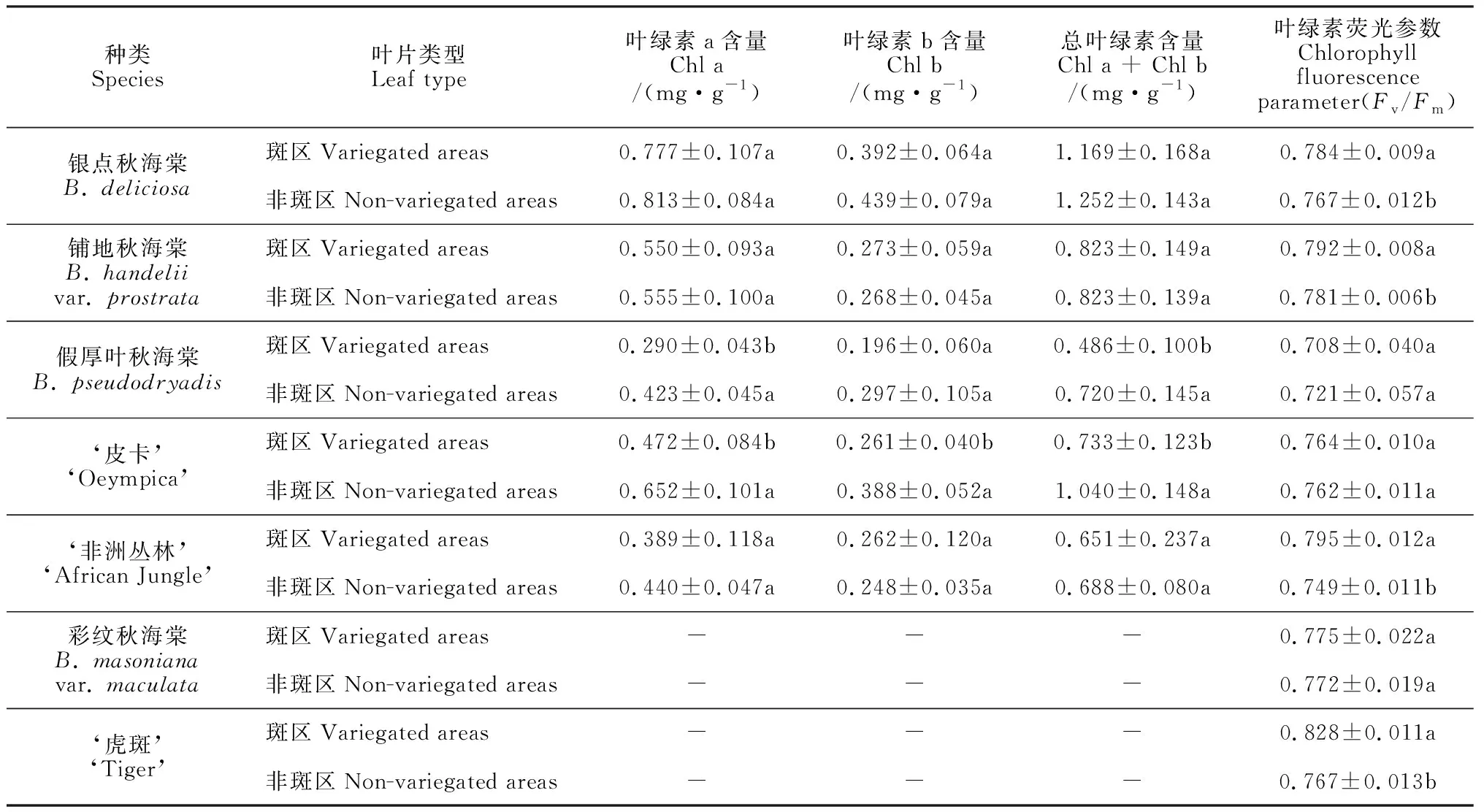
表1 7种秋海棠斑区和非斑区叶绿素含量及叶绿素荧光参数Fv/Fm值
注:同一种类中不同字母表示叶片类型间在0.05水平存在显著性差异
Note: The different normal letters within same species indicate significant difference among leaf types at 0.05 level

表2 4个组合杂交后代性状分离
3 讨 论
叶斑可分为2种类型:结构型(表皮型和空隙型)和色素型(叶绿素型和色素型),空隙结构型叶斑是由于空隙结构的存在使光线到达绿色组织时发生二次反射,在叶片表面表皮细胞边缘形成白色多边形的光反射而不是普通叶片表皮细胞中央形成的白色点状光反射,使该区域相对周围的正常叶片偏白,从而形成叶斑[12,16]。也有研究发现当叶背面颜色为红色时,与绿色相结合后,叶腹面会呈现深绿色,深绿色在有空隙结构存在的情况下呈现浅绿色,而不是白色[13,17]。本研究7种(品种)秋海棠的叶片半薄切片观察结果显示银点秋海棠、铺地秋海棠、假厚叶秋海棠、‘皮卡’和‘非洲丛林’叶片斑区的上表皮细胞与栅栏组织细胞间存在空隙,非斑区则没有空隙。所以他们的叶斑属于空隙结构型;彩纹秋海棠和‘虎斑’的斑区和非斑区上表皮细胞与栅栏组织细胞间均没有空隙,而斑区叶色为褐色或红褐色,其叶斑则属于色素型叶斑。
前人研究大王秋海棠(B.rexPutz.)[12]、水鸭脚秋海棠(B.formosana(Hayata) Masam.)、掌叶秋海棠(B.hemsleyanaHook. f.)、变色秋海棠(B.versicolorIrmsch.)、B.chlorostictaSands、B.diademaLinden、B.pustulataLiebm、B. ‘K030960’[13]和古林箐秋海棠(B.gulinqingensisS. H. Huang & Y. M. Shui)等[17]9种秋海棠叶斑形成时也发现了空隙结构,其中8种秋海棠叶斑的空隙结构存在于栅栏组织细胞与上表皮细胞间,栽培种B. ‘K030960’斑叶上表皮下方的储水组织细胞与栅栏组织细胞间存在空隙结构。另外,广西落檐[18]、仙客来[19]、猴枣猕猴桃[20]等具白色斑纹的叶片上表皮细胞与栅栏组织细胞间也存在空隙结构,这些具空隙结构型叶斑的种类其斑区的栅栏组织细胞与非斑区紧密排列的栅栏组织细胞不同,呈不规则漏斗状,排列较松散,含有少量的叶绿体。本研究中银点秋海棠、铺地秋海棠、假厚叶秋海棠、‘皮卡’和‘非洲丛林’5种具空隙结构的秋海棠斑区栅栏组织细胞也呈不规则漏斗状或呈不规则圆形,含有较少的叶绿体,植物叶片在植物进化过程中对环境变化比较敏感且可塑性较大,环境变化常导致叶的厚度、叶肉栅栏组织等形态解剖结构的响应与适应[21],由于具空隙结构型叶斑的种类包括秋海棠多生长于热带雨林的低层树林下,推测空隙型结构是为适应低光照环境而长期进化的结果,另外,也有研究认为这种空隙结构型叶斑是一种伪装色,避免受到食草动物的伤害。本研究还发现银点秋海棠和铺地秋海棠每个斑点中央有一个毛状体, Sheue等[13]在B.diadema和掌叶秋海棠、Chen 等[22]在柏拉木中也有类似发现,Chen 认为毛状体中含有晶体,可能与柏拉木空隙结构叶斑形成有关。
叶绿体超微结构显示银点秋海棠、铺地秋海棠、假厚叶秋海棠、‘皮卡’和‘非洲丛林’5种秋海棠斑区和非斑区形状基本一致,且均含有丰富的类囊体膜和密集的基粒片层,在斑区还观察到淀粉粒。Chen等[22]在研究柏拉木和蜂斗草空隙结构型叶斑时,也发现斑区含有密集的类囊体,基粒片层清晰,同时观察到淀粉粒。在叶绿素含量方面,银点秋海棠、假厚叶秋海棠、‘皮卡’和‘非洲丛林’斑区的叶绿素a、b及总叶绿素含量均低于非斑区,铺地秋海棠斑区叶绿素a、b及总叶绿素含量与非斑区差别不大,但5个种类斑区叶绿素荧光参数Fv/Fm值高于非斑区或与非斑区无显著差异,Rocca等[23]研究意大利魔芋光系统Ⅱ (PSⅡ) 活性时发现不同颜色叶片之间的最大光化学效率(Fv/Fm)无显著差异,更进一步说明空隙结构型叶斑不是由于叶绿素缺失造成,而且斑区的光能利用率更高。
目前关于植物叶斑的研究主要集中在色素型叶斑,尤其是针对叶绿素缺失造成的叶斑研究较多,对空隙结构型叶斑研究则相对较少,叶斑遗传特性方面的分析鲜见。本研究通过3个杂交组合和1个自交组合探讨空隙结构型叶斑的遗传特性,杂交、自交组合的后代均发生性状分离,产生有斑植株和无斑植株,其中自交组合获得的杂交后代株数最多,共获得387株,有斑和无斑植株比例接近3∶1,3个杂交组合中银点秋海棠(♀)×水鸭脚秋海棠(♂)获得338株杂交后代,银点秋海棠(♀)×长翅秋海棠(♂)后代共284株,光滑秋海棠(♀)×银点秋海棠(♂)后代205株,3个杂交组合中有斑和无斑植株的比例约为1∶1。 Henny[24-26]经过多年杂交试验发现粗肋草属植物空隙型叶斑是由单一位点复等位基因Vv控制,vv表现为叶片无斑,V-表现为有斑,不同的显性基因,斑纹位置和形状也不相同,不同显性基因同时存在时性状表现为共显性,至今已发现至少6个共显性等位基因:V、Vc、Vei、Vef、Vmn、Vtt。李景秀[27]通过有性杂交方法探讨毛叶秋海棠(B.rex)遗传杂交特性,发现毛叶秋海棠银绿色环状斑纹相对无斑为显性性状,其杂交遗传特性符合孟德尔分离定律。虽然空隙型结构叶斑多是由叶片上表皮细胞与栅栏组织细胞之间的空隙造成,但不同植物不同叶斑类型的遗传特性并不相同,结合前人研究,推测银点秋海棠叶片白色点状斑与无斑是1对可遗传的相对性状,白色点状斑是显性性状,无斑是隐性性状,通过有性育种可使后代获得斑点。
猜你喜欢
种子(2022年2期)2022-03-21
中国现代医药杂志(2020年10期)2020-12-14
作文评点报·作文素材小学版(2019年38期)2019-11-16
科学之友(2019年10期)2019-10-29
满族文学(2019年5期)2019-10-08
江苏大学学报(自然科学版)(2018年6期)2018-11-15
领导决策信息(2017年14期)2017-06-21
中成药(2017年5期)2017-06-13
现代检验医学杂志(2015年6期)2015-02-06
筑路机械与施工机械化(2014年8期)2014-03-01
