
转PcLIPH8基因水稻的构建及其相关表型分析
2019-01-04 05:09林法明李光豪高俊峰杜长青
西北植物学报 2018年11期
李 珅,王 珂,林法明,李光豪,高俊峰,杜长青
(河南农业大学 农学院,郑州450046)
木质素是重要的植物次生代谢产物之一。在高等植物体内,木质素是仅次于纤维素的第二位最丰富的大分子有机物[1-2]。木质素沉积在木质部导管及韧皮部纤维等组织中,参与调控植物生长发育、病虫害防御及环境适应性等过程[3-7]。然而,植物体内过多的木质素含量会对造纸等工业中纤维素与木质素的分离造成很大影响,成为造纸等产业的主要废物,严重污染环境[8-11]。另外,饲草植物中木质素的含量还影响牲畜的消化及营养吸收。因此在不影响植物正常生长发育的前提下,降低木质素含量或改变其组成,将为解决中国的环境污染问题及提高植物品质做出重要贡献。
木质素的降解酶系[12-16]主要分为3大类:木质素过氧化物酶 (lignin peroxidases, LIP)、锰过氧化物酶 (manganese perxidases, MnP) 和漆酶 (laccase)。黄孢原毛平革菌 (Phanerochaetechrysosporium) 被认为是白腐真菌中木质素降解能力最强的一种。一般认为黄孢原毛平革菌只产生LIP和MnP,不产生漆酶[17]。有研究表明,黄孢原毛平革菌的LIP基因家族至少含10个以上结构上紧密关联的基因,分别被命名为LIP-A~LIP-J,且每个基因还有不同形式的等位基因[18-19]。自1998年将一个编码黄孢原毛平革菌LIP的基因克隆入E.coli中成功获得表达以来,黄孢原毛平革菌木质素相关基因的异源表达研究发展迅速[20]。王海宽等[21]从黄孢原毛平革菌中克隆了LIPH8基因并将其导入甲醇毕赤酵母中进行表达,获得了表达高活力木质素过氧化物酶的酵母工程菌株,表明黄孢原毛平革菌中的这些编码LIP的基因在木质素降解方面可能发挥着非常重要的作用。然而,这些LIP基因在转基因植物中的功能却鲜有报道。
水稻既是一种重要的粮食作物,也是一种良好的研究基因功能的模式植物。本研究将黄孢原毛平革菌编码木质素过氧化物酶的PcLIPH8基因克隆至植物双元表达载体pCAMBIAL-1301,构建重组表达载体35S∷PcLIPH8并遗传转化水稻野生型品种Kitaake,分析其在水稻中的表达情况及对水稻木质素含量的影响,为确定该基因在降低植物木质素含量,改善植物品质方面的潜在功能奠定了一定的前期基础。
1 材料和方法
1.1 实验材料
入门载体质粒pMD-18-PcLIPH8,植物双元表达载体pCAMBIAL-1301,野生型水稻种子(Kitaake),均为河南省水稻生物学重点实验室保存。
1.2 研究方法
1.2.135S∷PcLIPH8植物双元表达载体的构建及转基因水稻株系获得利用双酶切法,分别对携带SacI和NcoI酶切位点的入门载体质粒pMD-18-PcLIPH8及植物双元表达载体pCAMBIAL-1301进行双酶切,胶回收酶切产物并利用T4DNA连接酶进行连接反应,转化DH5α感受态细胞,菌落PCR筛选阳性菌株并送上海生工测序验证。提取测序正确的阳性菌株的重组质粒35S∷PcLIPH8,电击法将重组质粒转化入根癌农杆菌EHA105,并利用农杆菌介导法转化野生型水稻品种Kitaake的胚性愈伤组织,经脱菌、30 mg/L潮霉素筛选、抗性愈伤分化、生根等过程,培养至成熟期单株收种。T3代纯合株系用于下一步的表型分析。
1.2.2转PcLIPH8基因水稻株系的分子鉴定利用Invitrogen公司的TRIzol试剂分别提取野生型和转基因水稻株系幼苗的总RNA。琼脂糖凝胶电泳检测后,选取质量完好的RNA用于下一步的逆转录。取1 μg总RNA,利用宝生(TaKaRa)公司反转录试剂盒反转录合成cDNA,-20 ℃冰箱保存,用于转基因株系的鉴定。设计PcLIPH8的特异性引物所用引物均见表1,对转基因水稻株系中的PcLIPH8表达量分析,以野生型水稻作为阴性对照。PCR扩增程序为:94 ℃预变性5 min,94 ℃变性30 s,56 ℃复性30 s,72 ℃延伸30 s,共35个循环。以水稻OsACTIN1基因作为PCR的内参,3次生物学重复。2-ΔCt法分析qRT-PCR试验数据。半定量RT-PCR产物在1%琼脂糖凝胶上电泳并用凝胶成像系统分析。
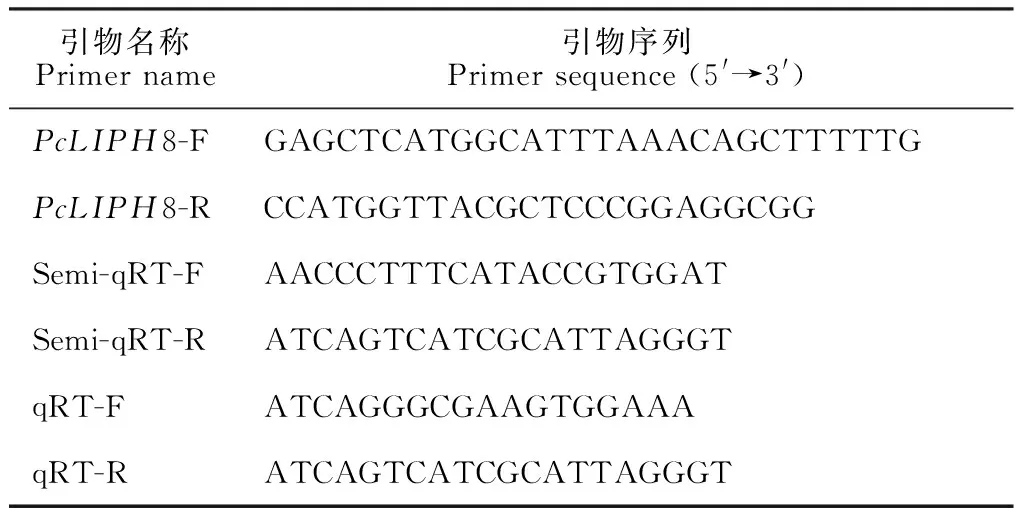
表1 引物列表
1.2.3木质素过氧化物酶的活性测定根据木质素过氧化物酶氧化藜芦醇生成藜芦醛,在310 nm处有特征吸收峰的特点,木质素过氧化物酶活性具体参照Tien等[22]的方法测定。取0.1 g新鲜水稻幼苗样品,在液氮中充分研磨,加入0.05 mol/L酒石酸钠缓冲液(pH2.5)1 mL匀浆,10 000 r/min、4 ℃离心10 min,取上清用于木质素过氧化物酶活性鉴定。反应体系为5 mL,含0.05 mol/L酒石酸钠缓冲液(pH 2.5),1 μmol/L藜芦醇,0.2 mmol/L过氧化氢,1 mL上清液。观察记录310 nm 处吸光值变化。每克鲜重样品每分钟氧化1 nmol 藜芦醇所需的酶量为一个酶活力单位(U)。
1.2.4木质素含量测定根据木质素中的酚羟基发生乙酰化后在280 nm处有特征吸收峰,且280 nm吸光值低与木质素含量正相关的原理[23],具体
参照Syros等[24-25]的方法测定。首先将样品80 ℃烘干至恒重,粉碎,过40 目筛,称取2 mg样品置于10 mL 玻璃试管中用于测定。 向干燥物中加入500 μL 25%溴乙酰 (冰醋酸配置) 和20 μL高氯酸,封口膜密封,充分混匀,80 ℃水浴40 min,每隔10 min 震荡1次,然后自然冷却至室温。加入500 μL NaOH-冰乙酸混合溶液,充分混匀,终止反应。5 000 r/min离心10 min,取上清20 μL,加冰乙酸稀释至1 mL。取200 μL于96孔板,测定280 nm处吸光值。以未加样品的作为空白对照。
1.2.5转PcLIPH8基因水稻株系的表型观察在成熟期对转PcLIPH8基因水稻株系进行表型观察分析,包括株高、节间长度、穗长等,拍照记录。试验数据以平均值±标准差表示,应用SPSS16.0统计软件分析差异显著性。
2 结果与分析
2.1 35S∷PcLIPH8植物双元表达载体构建及转基因水稻株系的获得
将重组阳性菌落进行菌落PCR鉴定及测序验证,结果表明成功构建了植物双元表达载体35S∷PcLIPH8 (图1,A)。将含有目的基因的重组表达载体电击法转化入根癌农杆菌EHA105中,遗传转化野生型水稻品种 (Kitaake) 获得潜在转基因植株。PCR鉴定阳性转化子结果表明,共获得3个独立的阳性转化子 (图1,B)。利用半定量RT-PCR及定量RT-PCR技术,对转基因水稻株系中PcLIPH8基因的转录水平进行检测,结果表明,在获得的3个转PcLIPH8基因独立转化子株系 (Ox-1、 Ox-3和Ox-6) 中PcLIPH8均有较高的表达量,而未转基因的野生型株系中未检测到该基因的表达,说明外源黄孢原毛平革菌的木质素过氧化物酶基因PcLIPH8在水稻中成功异源表达 (图1,C、D)。此外,我们观察到转PcLIPH8基因水稻与野生型相比,在苗期表型上并无显著性差异 (图1,E)。
A. 35S∷PcLIPH8示意图;B. PCR鉴定: 1~9 潜在转基因水稻植株;C、D. 半定量和定量RT-PCR;E. 苗期表型;WT. 野生型; Ox-1、Ox-3和Ox-6. 转基因株系图1 植物双元表达载体35S∷PcLIPH8的构建及转基因水稻株系的鉴定A. Schematic representation of 35S∷PcLIPH8; B. PCR confirmation: 1-9 potential transgenic rice lines; C and D. Semi-quantitative and quantitative RT-PCR analysis; E. Phenotype analysis at seedling stage; WT. Wild-type; Ox-1, Ox-3 and Ox-6. Transgenic rice linesFig.1 Generation of plant binary expression vector 35S∷PcLIPH8 and identification of transgenic rice lines
2.2 转PcLIPH8基因水稻株系的木质素过氧化物酶活分析
为了进一步证明PcLIPH8可以在水稻中异源表达,以10 d龄转PcLIPH8基因水稻幼苗为材料,提取植物总蛋白,进行木质素过氧化物酶活性测定。以未转基因10 d龄野生型水稻幼苗作为对照。结果表明,转PcLIPH8基因水稻幼苗的木质素过氧化物酶活性显著高于野生型水稻幼苗,增幅3.06%~5.07%,表明将PcLIPH8基因遗传转化入水稻中提高了其木质素过氧化物酶活性 (图2)。
2.3 转PcLIPH8基因水稻株系的木质素含量分析
以相同培养条件下的野生型水稻株系作为对照,进一步分析幼苗期及成熟期转PcLIPH8基因水稻的木质素含量变化。结果表明,无论是幼苗期还是成熟期,转PcLIPH8基因水稻幼苗的木质素含量均显著低于野生型水稻幼苗,苗期降低11.44%~14.97%,成熟期降低13.83%~20.05% (图3),表明将PcLIPH8基因异源表达入水稻中通过提高其木质素过氧化物酶活性有效降低了木质素含量。

**表示野生型与转基因株系之间差异极显著(P<0.01);下同图2 转PcLIPH8基因水稻株系的LIP活性分析** indicate significant differences at 0.01 level between wild-type and transgenic rice lines; The same as belowFig.2 LIP activity assay of transgenic PcLIPH8 rice lines
2.4 转PcLIPH8基因水稻株系的株高变化
对转PcLIPH8基因水稻植株的成熟期表型观察发现,转PcLIPH8基因水稻株系的株高显著高于未转基因的野生型水稻株系,株高较野生型增加约30% (图4,A、B)。对其穗长及各节间长度比较分析表明,转PcLIPH8基因水稻株系的穗长及Ⅰ、Ⅲ、Ⅳ节间均显著长于野生型对照,Ⅱ节间在转PcLIPH8基因水稻株系和野生型水稻株系中并无显著性差异 (图4,C、D),说明转PcLIPH8基因水稻株系株高的增加主要是由于穗长及Ⅰ、Ⅲ、Ⅳ节间的增长决定。
2.5 转PcLIPH8基因水稻株系的生物量变化
对成熟期转PcLIPH8基因水稻植株的生物量进行了相关表型观察分析,发现转PcLIPH8基因水稻株系的穗粒数显著高于野生型水稻,穗谷粒数增多110%~120% (图5,A~C),然而转PcLIPH8基因水稻株系的谷粒长度较野生型变短6.67%~7.15% (图5,D、E)。为分析这些农艺性状变化对产量及生物产量的影响,进一步比较了千粒重变化,转PcLIPH8基因水稻株系的千粒重降低,较野生型水稻减小12.86%~13.34% (图5,F), 而单株的生物量却增大18.61%~22.97% (图5,G)。表明PcLIPH8基因转入水稻虽然在一定程度上降低了千粒重,籽粒变小,但却极大地增加了穗粒数及生物量,为利用该基因改善农作物的生物产量奠定了一定的前期基础。
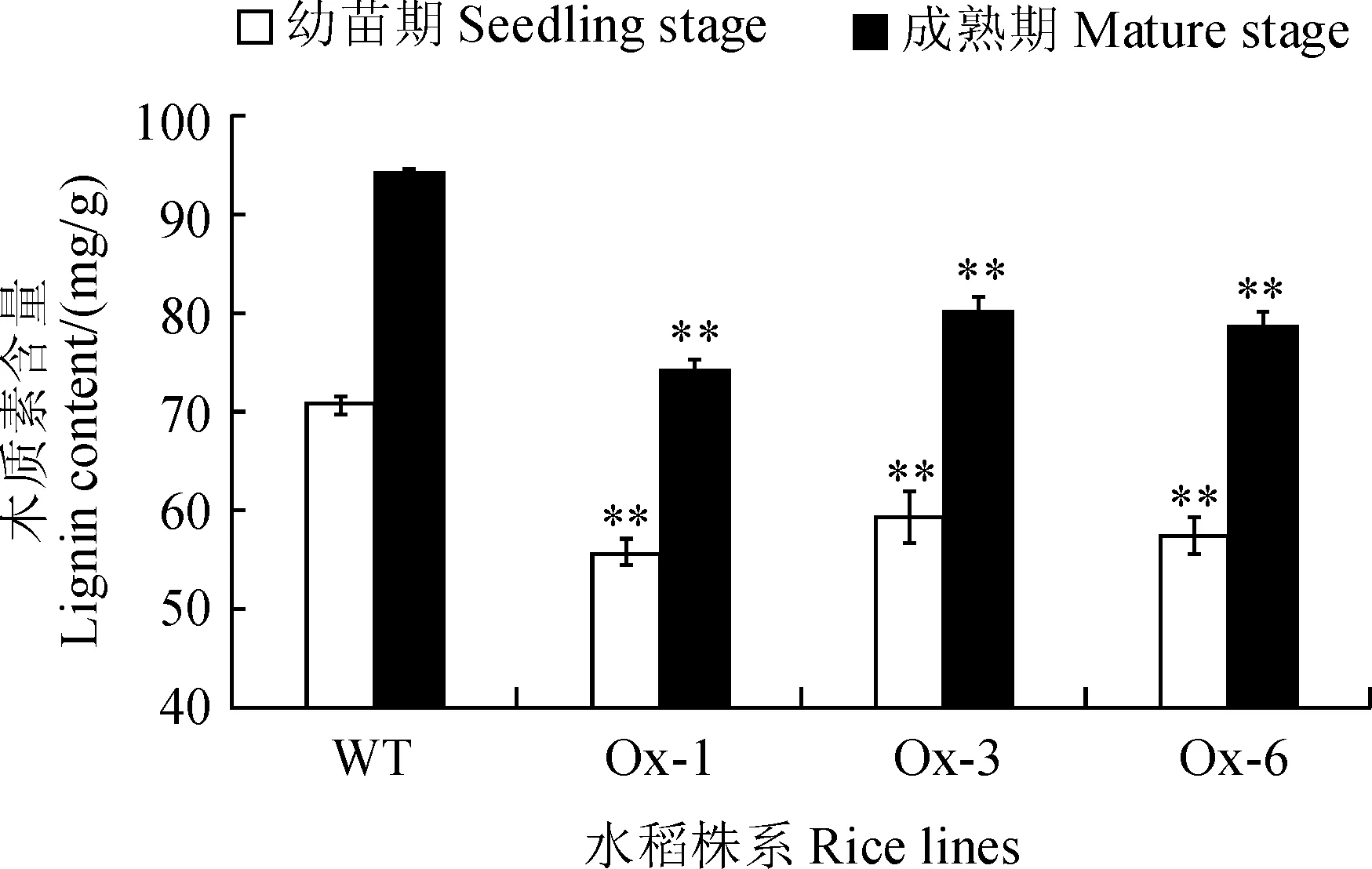
图3 不同时期转PcLIPH8水稻株系的木质素含量比较Fig.3 Lignin content of transgenic PcLIPH8 rice lines at different stages

A、B. 株高表型及统计分析, Bar=20 cm;C、D. 穗及节间长度表型及统计分析, Bar=10 cm. PL.株高;Ⅰ~Ⅳ. 第1~4节间图4 转PcLIPH8基因水稻株系的株高及节间长度分析A and B. Plant height phenotype and statistical analysis, Bar=20 cm;C and D. Panicle and internode length phenotype and statistical analysis, Bar=10 cm. PL. Plant height; Ⅰ-Ⅳ. The 1st, 2nd, 3rd and 4th internodeFig.4 Analysis of plant height and internode length of transgenic PcLIPH8 rice lines

A. 穗表型分析,Bar=5 cm;B、C. 穗粒数及统计分析,Bar=5 mm;D、E. 谷粒长度分析;F. 千粒重分析;G. 单株生物量分析图5 转PcLIPH8水稻株系的生物产量分析A. Panicle phenotype analysis, Bar=5 cm; B and C. Panicle grain number and statistical analysis, Bar=5 mm; D and E. Grain length analysis; F. Thousand grain weight analysis; G. Biological production analysisFig.5 Biological production analysis of transgenic PcLIPH8 rice lines
3 讨 论
木质素作为一种重要的次生代谢物,在植物生长、发育、抗病虫等方面具有十分重要的意义[26]。然而,过多的木质素以及不同的木质素结构组成大大制约了一些植物的利用效率及经济价值,因此在不影响植物正常生长发育的前提下,降低其木质素含量或改变其组成,已成为植物木质素研究的热点和重点。
随着现代植物生物技术的发展,转基因研究大大加快了对基因功能的揭示。研究表明将一些编码某些重要酶类的基因异源表达至高等植物体内可显著改善植物的某些表型。如杜长青等[27]将真菌Sclerotiniasclerotiorum的谷氨酸脱氢酶基因SsGDH在水稻中异源表达可增强对除草剂的抗性。因此,通过基因工程技术降低植物木质素含量可能是提高植物经济价值,减少环境污染的一条有效途径。钟天秀等[28]从华南象草中克隆了木质素合成关键酶基因Pp4CL并将其转入烟草,发现能显著提高转基因烟草的木质素含量。黄孢原毛平革菌是白腐真菌的模式菌种,对木质素具有极强的降解能力。黄孢原毛平革菌对木质素的强降解能力主要取决于其产生的2种酶:LIP和MnP。碳源、氮源、氧气、微量元素等外界因素均能显著影响黄孢原毛平革菌木质素降解酶的产生[29]。LIP是第一个从黄孢原毛平革菌中被发现的木质素降解酶,在木质素降解中发挥着关键作用。LIP基因排列紧密且高度保守,以基因家族形式存在。然而,这些LIP基因在转基因植物中的功能却并不十分清楚。
本研究将编码黄孢原毛平革菌木质素过氧化物酶的PcLIPH8基因导入野生型水稻品种Kitaake中异源表达,分析其对水稻木质素含量的影响及相关表型变化,结果表明将外源PcLIPH8基因导入水稻后,可显著提高转基因水稻的木质素过氧化物酶活性,增强其木质素降解能力,从而有效降低木质素含量。成熟期的田间表型分析表明,尽管导入外源PcLIPH8基因导致转基因水稻的千粒重在一定程度上降低,籽粒变小,但是转基因水稻株高显著变高,单株籽粒数增多,总生物量显著增加。此外,转PcLIPH8基因水稻的茎节较野生型水稻品种变脆变薄,可能会在一定程度上影响水稻的抗倒伏性。尽管如此,这些结果充分说明了外源PcLIPH8基因在降低植物木质素含量,提高生物产量等方面的潜在功能,为下一步利用该基因降低某些特定植物的木质素含量,提高其生物产量,从而提高其经济价值奠定了一定的前期基础。
猜你喜欢
河北科技师范学院学报(2022年2期)2022-08-26
亚热带植物科学(2022年1期)2022-05-17
生命科学研究(2021年1期)2021-01-25
上海包装(2019年8期)2019-11-11
四川农业科技(2019年5期)2019-07-01
分析化学(2018年7期)2018-09-17
天津造纸(2016年1期)2017-01-15
中国塑料(2016年3期)2016-06-15
浙江柑橘(2016年1期)2016-03-11
中国造纸学报(2015年1期)2015-12-16
