
丛枝菌根真菌(AMF)对星油藤根腐病的抗性研究
2019-01-04 06:05:40葛立傲普晓兰马焕成伍建榕
西南林业大学学报 2018年6期
唐 燕 葛立傲 普晓兰 马焕成 伍建榕
(1.西南林业大学国家林业局西南地区生物多样性保育重点实验室,云南 昆明 650224;2.西南林业大学云南省高校森林灾害预警控制重点实验室,云南 昆明 650224;3.上海出入境检验检疫局,上海 200000)
星油藤 (Plukenetiavolubilis) 又称印加果,是大戟科 (Euphorbiaceae) 的一种木质藤本植物,因果形为四角形、五角形或六角形,因此中国科学院西双版纳植物园将其命名为星油藤[1-2]。2006年,中国科学院院长路甬祥从秘鲁将星油藤引种到中国科学院西双版纳植物园,栽培成功。目前,星油藤已在云南省引种示范近70 hm2[3]。星油藤种子含有大量的不饱和脂肪酸、蛋白质,还有多种维生素和生育酚,故具有较高的研究价值[4-5]。但是,由于大量的人工种植,星油藤的病害日趋严重,星油藤的主要病害分为地上部分和地下部分。地上部分病害还可以通过物理或化学防治等措施进行防御,而地下部分病害主要为根腐病,既不能为人们预先发现,也难以控制,只有当星油藤叶片变色时才得以发现[6-8]。导致星油藤根腐病的病原菌具有多样性,不但真菌病原可以侵染,细菌也可侵染。据研究发现,星油藤根腐病的主要病原菌为尖孢镰刀菌 (Fusariumoxysporum)[9],已成为其最为严重的病害之一,也是影响星油藤产业化的重要障碍[3]。
菌根是土壤中的菌根真菌与高等植物的根系形成的一种共生体,是自然界中一种普遍的植物共生现象,大多数植物包括苔藓、蕨类、裸子植物、被子植物都能形成菌根[10]。菌根分为泡囊-丛枝菌根 (VAM/AM)、外生菌根 (ECM) 等7种类型[11]。丛枝菌根真菌 (AMF) 是广泛存在于土壤生态系统中的一类有益微生物,可以与陆地上90%以上的维管植物形成共生体[12-13]。研究发现,AMF与寄主植物形成共生关系后,能改善土壤理化特性、增加植物养分和水分的吸收与利用、增强植物抗逆性[14-15]、增加产量[16]还能能诱导植物对土传病原物产生抗病性[17-18]。研究表明,丛枝菌根真菌能够减轻一些土传病原真菌、孢囊线虫、根结线虫对植物造成的危害[19-20];另外,丛枝菌根真菌虽不全部都可以产生泡囊,但可以全部产生丛枝,改变植物根系的结构,当病原菌侵入植物根系后,丛枝菌根真菌共生体可以促进植物根系增长、增粗,使根尖表皮加厚、细胞层数增多,从而促进植物的抗病性,减缓病原体侵染植物的过程[21]。
云南省西双版纳傣族自治州是我国星油藤主要的种植区,也是星油藤根腐病的主要发生区,然而西双版纳有我国唯一的热带雨林自然保护区,因此对病虫害不能采用化学防治措施,最好通过生物防治的方法来探讨星油藤的病虫害防治,以保证西双版纳的森林健康和生态安全。本研究主要以西双版纳星油藤人工栽培区为研究地点,调查星油藤根围丛枝菌根真菌孢子的密度以及根系侵染率对星油藤根腐病发病率和发病程度的关系,为星油藤的病害防治提供参考。
1 材料与方法
1.1 调查地点
本研究在2015年11月至2016年5月在云南省西双版纳傣族自治州开展,该地地处东经99°56′~101°50′,北纬21°08′~22°36′,多为中低山和丘陵区。
1.2 发病率和病情指数调查
对西双版纳星油藤根腐病进行发病率调查、病情指数调查以及根系和根际土壤的采集,用自封袋密封带回实验室进行孢子的筛选以及菌根的侵染检测。分别在云南省西双版纳傣族自治州东部、西部、南部、北部以及中部地区5个星油藤种植地块进行样本采集,每个采样地的发病地块与健康地块分别随机选取15株星油藤,共150株,然后对150株不同发病程度的星油藤和不发病地块星油藤进行地上部分调查和地下部分采样。地下部分采样包括星油藤的须根系及其周围的根际土壤。
采用赵仕光等方法对星油藤的根腐病病害进行分级[22],就是通过观察星油藤根部的腐烂情况与生长长势情况进行分级 (表1),计算病情指数和发病率。

(1)
发病率=发病株数×调查总株×100%

(2)
1.3 丛枝菌根真菌孢子测定
1.3.1丛枝菌根真菌孢子的分离
在所有健康地块和发病地块样品中分别随机选取25株土样,对上述每株星油藤的根际土壤进行湿筛沉淀法分离丛枝菌根真菌孢子,首先去除表层1 cm厚的杂物,挖4~20 cm深的土壤剖面,收集根际土壤约2 kg,装入自封袋带回实验室中让其自然风干后于常温下保存,备用。
将采集的根际土壤用湿筛沉淀法进行处理,首先称取风干的根际土壤20 g,加入无菌水500 mL,搅拌,浸泡过夜。过夜后的土壤悬浮液分别通过20、100、140、200、300目的筛子,反复用无菌水进行冲洗[23]。再分别通过加入滤纸的布式漏斗过滤,在体视显微镜下分别统计4个筛面上的孢子。
1.3.2丛枝菌根真菌孢子密度的计算
将统计过的4个筛面的孢子数进行相加再除以20,得到每个土样的孢子密度。
1.3.3丛枝菌根真菌孢子的鉴定
在体视显微镜下,利用解剖针将单个孢子挑取到载玻片上,在显微镜下观察单个孢子的颜色、大小;将单个孢子压破,观察孢壁的层数、厚度、压迫的开口类型等。利用显微拍照技术对单个孢子进行拍照,利用 《VA菌根真菌鉴定手册》 和西弗吉利亚大学 (http://invam.caf.wvu.edu) 的网站上所提供的孢子形态,对孢子进行鉴定[24-25]。
1.4 丛枝菌根真菌对星油藤根系的侵染率测定
1.4.1根样的采集
对星油藤进行采集主根系和须根系,将根系剪成2 cm放入加有1/2的FAA (70%的乙醇90 mL、福尔马林5 mL、冰醋酸5 mL,稀释1倍) 的青霉素小瓶中,放入4 ℃的冰箱中保存。
1.4.2星油藤根系的透明染色
将根样从FAA中取出,用无菌水进行冲洗干净 (4~5次为宜),放入15 mm × 15 mm的试管中,加入15%的KOH,水浴2 h进行解离,从水浴锅中取出,自然冷却,用无菌水清洗4~5次,用2%的乳酸进行酸化20 min,清洗,加入0.05%的苯胺蓝 (乳酸333 mL、甘油333 mL、蒸馏水333 mL、苯胺蓝0.5 g) 进行染色,水浴1 h,用乳酸进行洗净。在光学显微镜下观察丛枝菌根真菌的侵染情况。利用显微拍照技术进行拍照,统计计算侵染率[24-26]。
1.4.3丛枝菌根真菌侵染率计算

(3)
2 结果与分析
2.1 根系的侵染率与星油藤根腐病发病率的关系
通过对星油藤根系的透明染色,可以观察到几乎每一根段均有菌根真菌的侵染,但侵染情况不定。在被侵染的根段中可以看到丛枝菌根真菌的2种典型结构:一种是菌丝无隔膜,呈二叉分枝状的丛枝 (图1a);另一种是菌丝较粗,具有明显的泡囊结构 (图1b)。
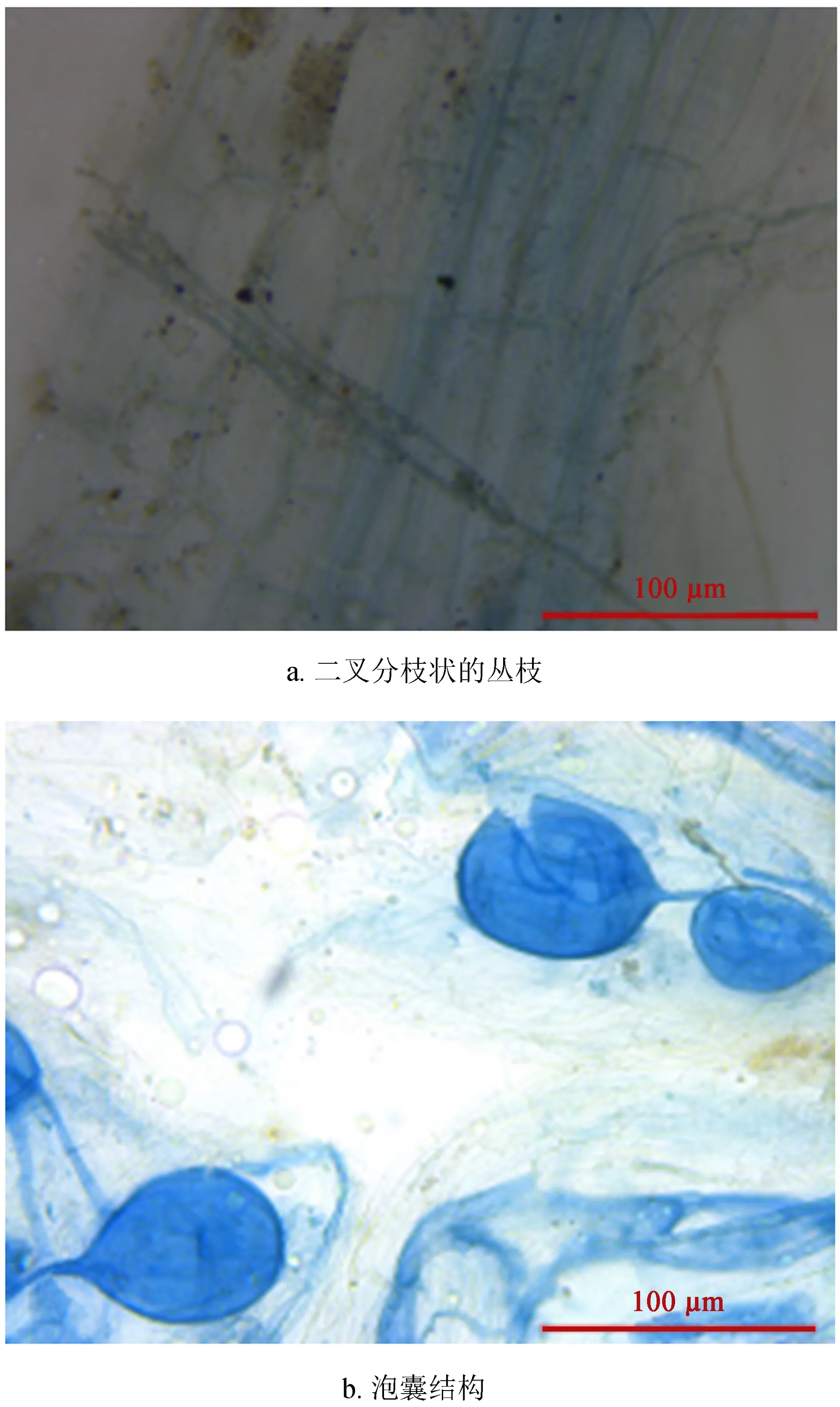
图1根系的透明染色
Fig.1 The transparent color of root
依据表2计算病情指数发现,调查的150株的发病指数达到43。在33株无病正常的星油藤中发现大量的丛枝和泡囊,菌丝的侵染率达到90%。而21株地上部分整株叶枯近死亡的星油藤中菌丝的侵染率只有13%,基本没有发现泡囊和丛枝的存在。
2.2 菌根真菌孢子密度与星油藤根腐病发病率的关系
通过表3发现,正常星油藤的孢子密度明显高于发病的星油藤孢子密度,而且随着病级的增加,孢子密度呈现减少的趋势。当孢子密度小于3时,植株病级达到Ⅴ级,星油藤基本不具有抗病性,基本接近死亡。当孢子密度大于6时,少数星油藤病级为Ⅰ级,但也能生长,其余大多数为0级,是正常植株。
通过计算发病率得到发病地块与健康地块星油藤根部丛枝菌根真菌孢子密度与根部病害发病率的关系。在发病地块中,当孢子密度低于2时,星油藤发病率达到60%,且发病严重。在健康地块中,丛枝菌根真菌孢子密度达到6时,发病率仅为4%,星油藤可以健康生长。
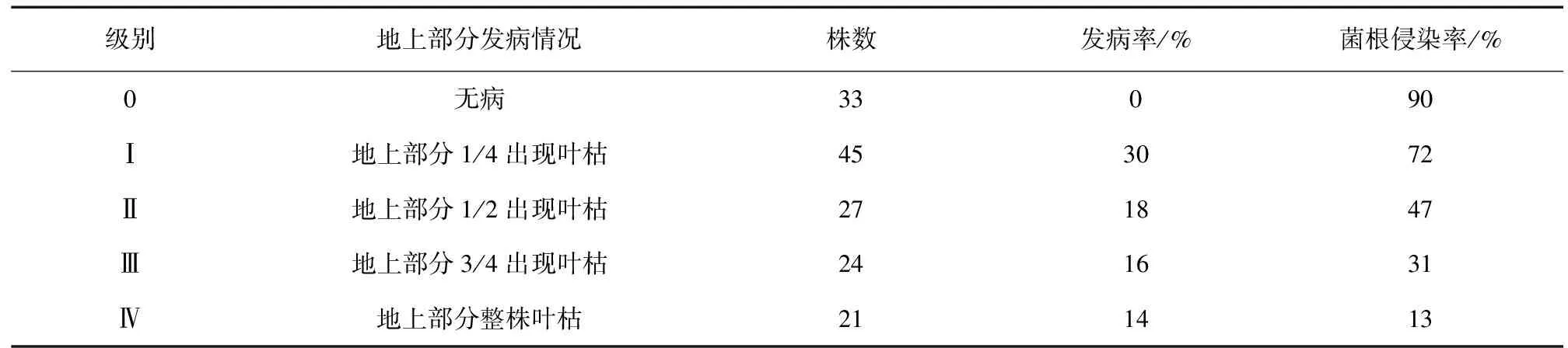
表2 发病情况、发病率及菌根侵染率Table 2 The morbidity,incidence of root rot and mycorrhizal infection rate
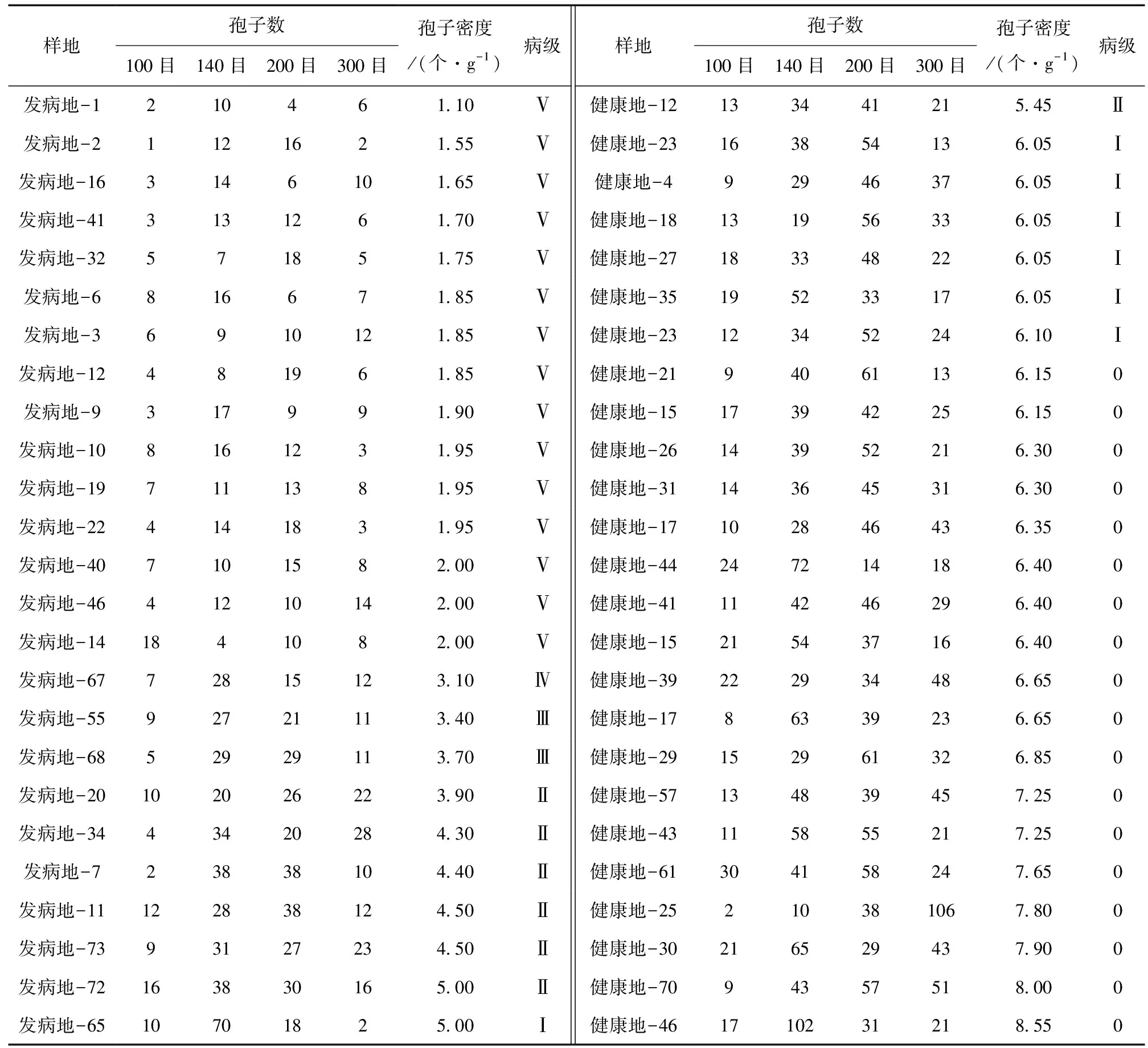
表3 孢子密度与病级统计Table 3 The statistics of spore density and disease degree
2.3 丛枝菌根真菌孢子的种类
通过星油藤土壤孢子的分离,共分离出1属3种AMF,分别为摩西球囊霉 (Glomusmosseae) (图2a)、幼套球囊霉 (Glomusetunicatum) (图2b) 和缩球囊霉 (Glomusconstrictum) (图2c)。
摩西球囊霉:幼嫩的孢子乳白色,成熟时浅黄—黄褐色,表面光滑,直径110~290 μm。孢壁3层,厚2~7 μm,孢子壁1层 (W1) 无色透明, Melzer′s试剂中染成粉红色;表面时常脱落形成颗粒状,在成熟的孢子中脱落;孢子壁2层 (W2) 无色透明,不易观察;孢子壁3层 (W3) 浅黄、黄褐色,层积壁。孢子内含物为大小不等的油滴或颗粒。连孢菌丝漏斗形,连点宽8~25 μm,连点处菌丝壁稍增厚;连孢菌丝底部漏斗形,有厚弯隔,是该种孢子的特征性结构[27]。
幼套球囊霉:孢子圆形或近圆形,表面光滑,黄色、黄褐色,直径70~120 μm。孢壁2层,壁厚4~13 μm。外层W1无色透明,易逝壁;内壁W2为层状壁,黄褐色。成熟孢子通常不见外层壁。连孢菌丝大多一条,无色易断,孢壁延伸入连孢菌丝一段距离,基部增厚至3~3.5 μm,连点宽7.5~12 μm,连点孔由隔封闭或由壁增厚封闭。内含物颗粒状[27]。
缩球囊霉:孢子球形或近球形,深红棕或黑棕色,孢壁光滑透亮,有时附着有碎屑,孢子直径80~160 μm。孢壁1层,为层状壁,厚6~12 μm,深黄棕、深红棕或黑棕色。连孢菌丝在连点处缢缩,宽11~15 μm,连点处壁增厚,封闭孔口或仅留一狭小通道,连点下方连孢菌丝膨大至17~19 μm宽,连孢菌丝颜色自连点处向下由黑红棕逐渐变浅至浅黄棕至浅黄。连孢菌丝通常向一侧弯曲,有时在连孢菌丝变宽部位下方有一隔,隔下有细分枝[27]。

图2丛枝菌根真菌孢子种类
Fig.2 The spore types of arbuscular mycorrhizal fungi
经过统计发现,在分离15株接近死亡的星油藤土壤中,只发现单属单种,并不具有丛枝菌根真菌的多样性。在18株健康的星油藤土壤中,分离出1属3种,并且,幼套球囊霉 (G.etunicatum) 是该株的优势种。
3 结论与讨论
3.1 丛枝菌根真菌菌根侵染率和孢子密度与根腐病的关系
星油藤根腐病是由20多种土传病原真菌、细菌复合侵染导致,发病植株生长发育受阻、产量和品质都受到严重的影响,已经成为制约星油藤产量发展的主要障碍。而丛枝菌根真菌可与星油藤共生,提高星油藤的抗病性。本研究发现,正常的星油藤根中有大量的丛枝和泡囊,同时菌丝的侵染率很高,发病的星油藤根中基本没有泡囊和丛枝的存在,同时菌丝的侵染率也低;并且正常星油藤的孢子密度明显高于发病的星油藤孢子密度,而且随着病级的增加,孢子密度减少。以上说明了丛枝菌根真菌与星油藤的抗根腐病有着密切的关系。但目前还未发现前人利用丛枝菌根真菌与星油藤共生达到抵抗根腐病的研究,但是有研究观察到健康三七 (Panaxnotoginseng) 的AMF侵染率明显高于有根腐病的三七,表明AMF提高了三七对根腐病的抗病能力[28]。也有学者发现AMF可通过促进植物养分吸收、生长,调控病害相关酶活性,提高紫花苜蓿对烟色织孢霉 (Microdochiumtabacinum) 引致的根腐病抗性[29]。
3.2 丛枝菌根真菌孢子的优势种与多样性与星油藤根腐病的关系
丛枝菌根真菌的优势种、多样性与星油藤根腐病也有一定的关系,研究表明:根系中具有丛枝菌根优势种和多样性较为丰富的星油藤抵抗根腐病的能力越强,发病率越低。由此可见,丛枝菌根真菌的多样性可提高星油藤的抗病性,减少其发病率。也有学者观察到对加工番茄根系接种摩西球囊霉 (G.mosseae) 和根内球囊霉 (G.intrara-dices) 菌种后,其根系侵染率分别为60.50%和47.50%,对加工番茄根腐病的防效为63.42%和21.22%[30]。Thygesen等[31]也用根内球囊霉 (G.in-traradices),近明球囊霉 (G.claroideum) 分别接种豌豆,发现2种AMF都能减轻豌豆根腐病病害。所以我们可以通过接种丛枝菌根真菌来预防星油藤根腐病的发生,提高星油藤的抗病性。
3.3 丛枝菌根真菌与病原菌相互作用
从丛枝菌根真菌在对健康地块植株和发病地块植株的侵染率来看,病原菌的存在降低丛枝菌根真菌的侵染率。原因可能是病原菌的侵害导致植物的生长长势衰弱,对丛枝菌根真菌的营养供应减少,丛枝菌根真菌生长减缓,导致侵染率下降。丛枝菌根真菌与病原菌都是异养型生物,都是依靠寄主植物提供营养才得以生存。而丛枝菌根真菌的存在必然会减少病原菌的营养供应,会对病原菌的生长起到一定的限制作用。
综合分析可知,丛枝菌根真菌的孢子密度、丛枝菌根真菌的多样性势必会提高星油藤的抗病性,促进星油藤根系的发育和营养生长。而丛枝菌根真菌诱导植物抗病性是一个影响植物根系的结果而影响植物功能和植物生理生化的复杂过程,包括影响根系周围土壤微生物的组成、病原物侵染植物的侵染位点等多个方面。并且不同栽培模式对油茶 (Camelliaoleifera) 土壤微生物及酶活性的影响也不一样,对它新梢生长、单株产果量及出油率也有显著影响[32],所以同样是油料植物的星油藤,不同栽培模式是否会影响土壤根系微生物的组成,从而影响丛枝菌根真菌种类的多样性以及植物的抗病性。而丛枝菌根真菌具体通过哪种机制提高植物的抗病性、以及这些机制的相互关系,需要进一步的探究。只有深入了解这些机制以及内在的联系,才能成功的将生物防治运用到植物的生产中来。
猜你喜欢
今日农业(2022年15期)2022-09-20 06:54:16
今日农业(2021年21期)2021-11-26 05:07:00
现代畜牧科技(2021年9期)2021-10-13 06:38:54
园林科技(2020年2期)2020-01-18 03:28:26
中成药(2018年3期)2018-05-07 13:34:24
上海蔬菜(2016年5期)2016-02-28 13:18:10
环境与可持续发展(2013年6期)2013-03-11 16:21:51
植物营养与肥料学报(2011年5期)2011-11-06 07:31:00
植物营养与肥料学报(2011年6期)2011-10-24 05:53:04
植物营养与肥料学报(2010年3期)2010-11-16 00:59:28
