
不同茎干类型云南松子代2年生苗木生长比较
2019-01-04 06:05陈永丽蔡年辉王大玮何承忠许玉兰
西南林业大学学报 2018年6期
张 焕 陈永丽 蔡年辉, 陈 诗 王大玮 何承忠 许玉兰
(1.西南林业大学西南山地森林资源保育与利用教育部重点实验室,云南 昆明 650224;2.西南林业大学云南省高校林木遗传改良与繁育重点实验室,云南 昆明 605224;3.商丘工学院园林教研室,河南 商丘 476000)
云南松 (Pinusyunnanensis) 是我国西南地区的特有树种,以云南省为其分布中心,向四川西部及西南部、贵州西部及西南部、西藏东部、广西西北部延伸分布[1];其适应性较强,耐干旱瘠薄,多为分布区的荒山造林先锋树种,在分布区域的国民经济建设和生态建设中发挥着非常重要的作用[2-3]。大量研究表明,除云南松天然更新幼林苗木或幼林地上部分生长缓慢,存在严重的 “蹲苗” 现象外[1,4],云南松林分也因人为逆向选择,导致优良种质资源减少严重,表现出一定的衰退现象,其中以茎干出现扭曲、弯曲现象等不良个体比例的逐渐增加较为显著[5],树干扭转会极大地影响林木的生长、木材的利用[6]和经济价值[7-8]。除此之外,林木生长周期长、林木基因杂合度高,自身和环境因素会影响其萌发、生长、开花及休眠,使得林木的育种周期延长,但林木早期优势的遗传现象是客观存在的。幼年时期的树木能够提供成年树木的生长表现,加之因种源和家系的遗传适应性的差异,从苗期就表现出其生长的分化,目前对云南松不同茎干类型子代苗期的研究不多。鉴于此,本研究以采自滇中云南松主分布区的玉溪市新平县、楚雄州禄丰县、昆明市宜良县3个地点的2种茎干类型云南松母株上的种子为试验材料,播种育苗,对2年生云南松苗木的苗高、地径、分枝高度及针叶长度等生长量指标进行分析,比较2种不同茎干类型云南松子代苗木的生长变异规律,以期为云南松种质资源的评价、保护、更新等提供基础信息和实践指导。
1 材料与方法
1.1 试验地概况
试验地设在西南林业大学苗圃内,位于昆明市北部,地处北纬25°04′00″、东经102°45′41″,海拔1 945 m。该地年均气温14.5 ℃,最热月 (7月) 平均气温19.7 ℃,最冷月 (1月) 平均气温7.5 ℃,年温差12~13 ℃;全年降雨量约1 031 mm、相对湿度74%,全年无霜期平均在240 d以上,晴天比较多,日照数年均2 445.6 h,日照率56%;终年太阳投射角度大,年均总辐射量达542.48 kJ/cm2。属于亚热带季风气候,干湿季分明,5—10月为雨季,降水量占全年的85%左右,11月至翌年4月为干季,降水量仅占全年的15%左右。
1.2 试验设计
1.2.1种子来源
种子来源为云南省昆明市宜良县、玉溪市新平县、楚雄州禄丰县3个采样地点的2种不同茎干类型的单株,单株间距为5倍树高以上,且为随机选取树冠中部成熟的饱满球果。种子采集地点的基本概况见表1。

表1 云南松采样地点基本概况Table 1 Basic information of P.yunnanensis sampling site
1.2.2苗木培育
试验采用完全随机区组设计,于2015年3月按采种地点分单株播种于西南林业大学格林温室大棚内。每个采种地点的种子播种50~60个家系,每个家系分别以5 cm × 10 cm的株行距播种2行,每行播20粒,共40粒种子,重复3次,每个家系播种120粒,发芽后进行常规的苗期管理,并对其苗期的生长状况进行观测和分析。
1.3 苗木测定
于2016年12月底云南松苗木生长停止时,从3个采样地点中随机抽取通直、弯扭类型的云南松子代苗木各10个家系 (共60个家系),对每个家系所有苗木的苗高、地径、分枝高度及针叶长度进行测定。苗高、分枝高度、针叶长度直接利用直尺测定,地径利用电子游标卡尺测定。
1.4 统计分析方法
利用Excel 2010对所测定的数据进行整理和图形的绘制;利用SPSS 21.0软件进行单因素分析和t检验,其中个别缺失数据按缺失处理。用变异系数 (CV) 来衡量表型性状的离散程度。即变异系数越大,离散程度越高。其计算公式为:

(1)

2 结果与分析
2.1 2种不同茎干类型云南松子代苗木的生长情况
通过对来源于3个采样地点的2种茎干类型云南松2年生子代苗木的生长量指标进行分析可知 (表2),2种茎干类型的云南松子代苗木的苗高、地径、分枝高度及针叶长度的平均值存在差异。通直类型的平均苗高、平均地径及平均分枝高度 (12.63 cm,11.94 mm,4.30 cm) 均稍小于弯扭类型 (13.88 cm,11.99 mm,5.33 cm),但在针叶长度方面,通直类型与弯扭类型间均值差异不大,且通直类型针叶长度均值 (22.77 cm) 略大于弯扭类型 (22.70 cm)。其中,苗高、分枝高度表现较高的是新平的弯扭类型,平均苗高为16.16 cm,平均分枝高度为13.63 cm;地径、针叶长度表现较高的是禄丰的弯扭类型云南松2年生子代苗木,平均地径为13.62 mm,平均针叶长度为23.20 cm。
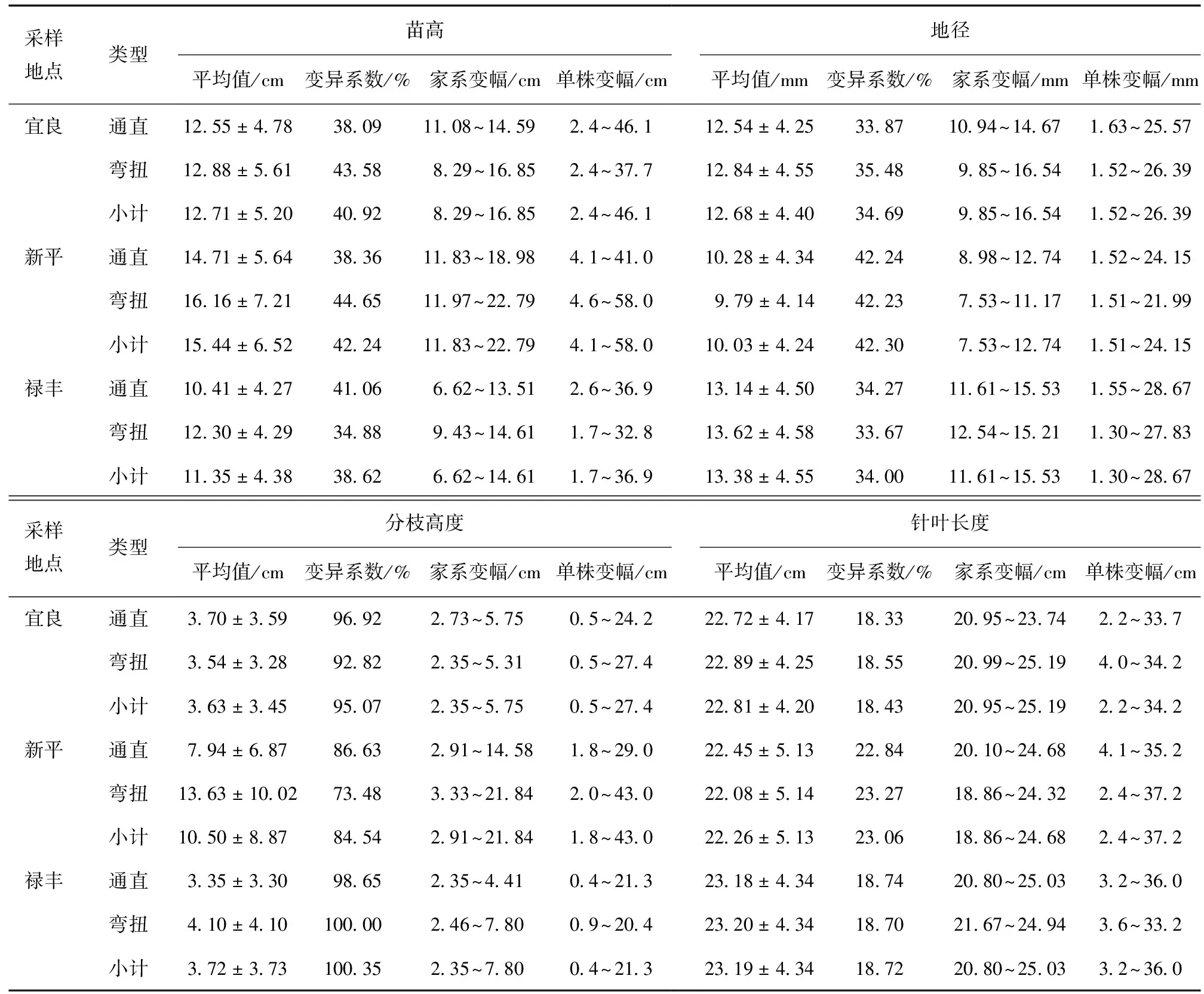
表2 两种不同茎干类型的云南松子代苗木的生长情况的统计分析Table 2 The statistical analysis of the growth of P.yunnanensis seedlings with 2 different stem types
从变异幅度方面来说,3个采样地点2种茎干类型云南松子代苗木的差异情况有所不同。宜良采样地点,除通直类型子代苗木的分枝高度均值极差 (3.02 cm) 略大于弯扭类型 (2.96 cm) 外,其苗高、地径、针叶长度的均值极差 (3.51 cm、3.73 mm、2.79 cm) 均小于弯扭类型 (8.56 cm、6.69 mm、4.20 cm)。新平采样地点2种类型的表现有些不同,除通直类型地径的均值极差 (3.76 mm) 稍大于弯扭类型 (3.64 mm) 外,其通直类型苗高、分枝高度及针叶长度的均值极差 (7.15 cm、11.67 cm、4.58 cm) 均小于弯扭类型 (10.82、18.51、5.46 cm)。禄丰采样地点各性状的均值极差却与宜良的恰恰相反,除分枝高度表现为弯扭类型 (5.34 cm) 大于通直类型 (2.06 cm) 外,其苗高、地径及针叶长度均为通直类型 (6.89 cm、3.92 mm、4.23 cm) 大于弯扭类型 (5.18 cm、2.67 mm、3.27 cm)。说明2种不同茎干类型的云南松子代苗木间的生长状况没有明显的区别,不同采样地点4个生长量指标的生长状况也不尽相同。
就单株极差而言,宜良采样地点的除地径与其均值极差表现类似外,其他指标生长表现与其均值极差相反,苗高、针叶长度的单株极差可达 (43.7、31.5 cm),弯扭类型的稍低但也达到 (35.3、30.2 cm),地径、分枝高度的单株极差是表现为通直类型 (23.94 mm、23.7 cm) 低于弯扭类型 (24.87 mm、26.9 cm);新平采样地点2种类型4个生长量指标的表现与宜良的不同,但其生长状况与均值极差相同,即除地径的单株极差表现为通直类型 (22.63 mm) 高于弯扭类型 (20.48 mm) 外,其通直类型的苗高、分枝高度及针叶长度的单株极差(36.9、27.2、31.1 cm)均小于弯扭类型 (53.4、41.0、34.85 cm);禄丰采样地点的苗高、地径、分枝高度及针叶长度4个生长量指标的单株极差均表现为通直类型的单株极差 (34.3 cm、27.12 mm,20.9 cm、32.8 cm) 大于弯扭类型 (31.1 cm、26.53 mm,19.5 cm、29.6 cm),与其均值极差相比,除分枝高度外均相似。由此表明,一方面云南松具有遗传多样性,存在着丰富的变异特性;另一方面云南松不同茎干类型子代苗木间差异不明显,各个采样地点间未表现出一致规律。
2.2 2种不同茎干类型的云南松子代苗木的变异系数分析
对2种不同茎干类型云南松子代苗木生长性状的变异进行分析,结果见图1~3,云南松不同生长生长性状变异系数存在较大的波动,变异范围为11.65%~128.89%。不同性状在不同类型间的变异系数也存在一定的波动,尤以分枝高度的变异系数波动最大。苗高、地径、分枝高度及针叶长度的变异系数在3个地点的平均表型变异系数分别为43.35%、38.69%、114.27%、20.28%,且通直类型苗高、地径、分枝高度、针叶长度的变异系数 (41.60%、37.96%、105.90%、20.13%) 均小于弯扭类型的变异系数 (44.24%、39.41%、118.12%、20.43%)。说明2种茎干类型均存在着极为丰富的遗传变异,但通直类型云南松子代苗木的生长性状比弯扭类型稳定。
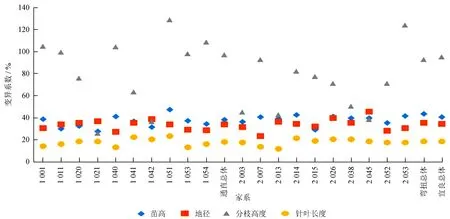
家系代号中首数字为1的是通直类型;首数字为2的是弯扭类型。
图1宜良云南松子代苗木变异系数分析
Fig.1 The variation coefficient analysis ofP.yunnanensisseedlings in Yiliang

家系代号中首数字为1的是通直类型;首数字为2的是弯扭类型。
图2新平云南松子代苗木变异系数分析
Fig.2 The variation coefficient analysis ofP.yunnanensisseedlings in Xinping

家系代号中首数字为1的是通直类型;首数字为2的是弯扭类型。
图3禄丰云南松子代苗木变异系数分析
Fig.3 The variation coefficient analysis ofP.yunnanensisseedlings in Lufeng
在同一类型内,不同性状在不同采样地点间的变异情况也有差异,但差异不大。对通直类型的云南松子代苗木来说,禄丰的苗高、分枝高度的变异系数最大 (41.06%、98.65%),新平的地径、针叶长度的变异系数最大 (42.24%、22.84%),宜良的4个生长量指标的变异系数处于中间位置,进而可以表明其离散程度较小;对于弯扭类型的云南松子代苗木来说,除新平苗高的变异系数 (44.65%) 为最大外,其他3个生长量指标与通直类型相似,即新平的地径、针叶长度的变异系数 (42.23%、23.27%)、禄丰的分枝高度的变异系数 (100.00%) 为最大,同样,宜良采样地点弯扭类型的变异处于中间水平。表明在不同茎干类型云南松子代苗木的生长过程中,弯扭类型的各生长量指标性状不整齐,较通直类型具有更高的变化幅度。
2.3 2种不同茎干类型的云南松子代苗木的方差分析
对2种不同茎干类型的云南松子代苗木进方差分析,结果见表3~4。3个采样地点间同一类型内云南松子代苗木的苗高、地径、分枝高度及针叶长度4个生长量指标进行单因素分析结果显示 (表3):同一类型内不同采样地点间云南松子代苗木的4个生长量指标差异性表现不同,但均达到差异极显著水平。
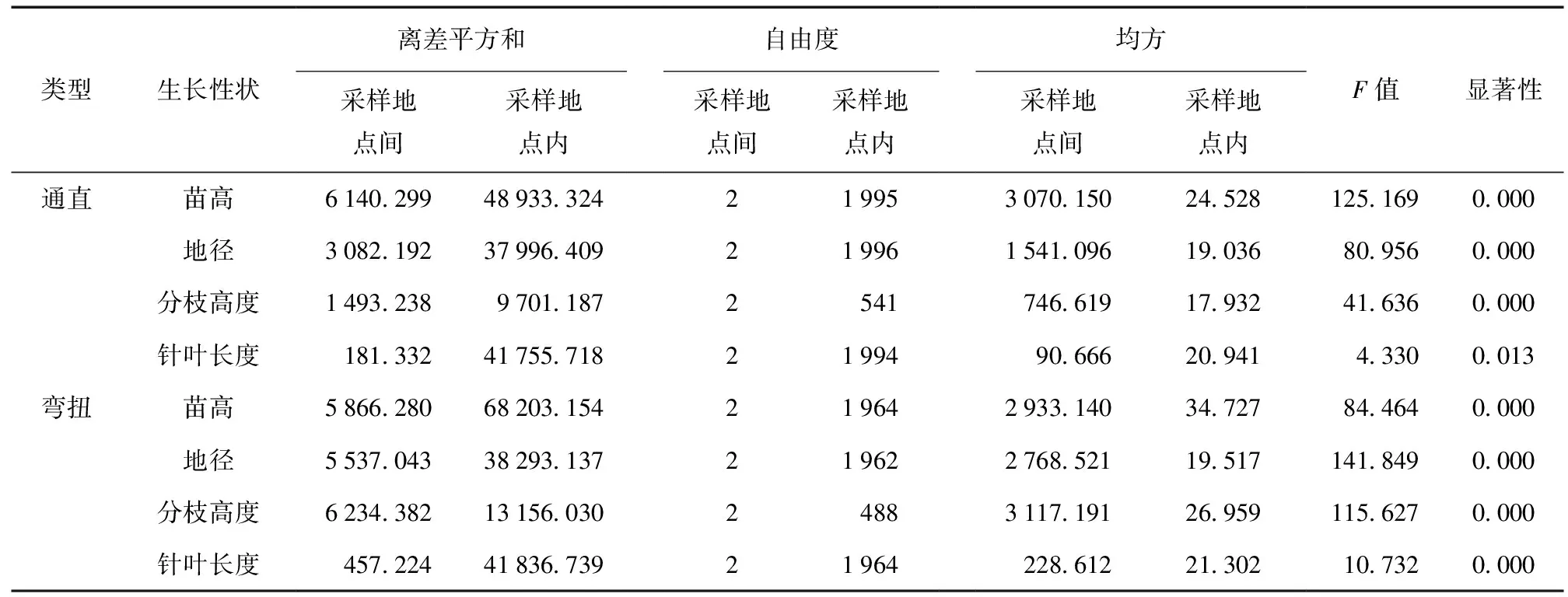
表3 不同采样地点间云南松子代苗木方差分析Table 3 The variance analysis of P.yunnanensis seedlings in different sampling sites

表4 两种不同茎干类型云南松子代苗木t检验Table 4 The t test of P.yunnanensis seedlings with 2 different stem types
对3个采样地点的2种茎干类型云南松子代苗木的苗高、地径、分枝高度及针叶长度4个生长量指标进行t检验分析可知 (如表4),宜良采样地点2种茎干类型的4个生长量指标间的差异性均为不显著 (P> 0.05),且通直类型苗高、地径、针叶长度的均值小于弯扭类型;新平采样地点的苗高、分枝高度为极显著差异 (P< 0.01),地径为显著差异 (P< 0.05),针叶长度为不显著差异 (P> 0.05),且弯扭类型地径及针叶长度的均值小于通直类型;禄丰采样地点的苗高为极显著差异 (P< 0.01),分枝高度为显著差异 (P< 0.05),而地径和针叶长度间的差异不显著 (P> 0.05),且4个生长量指标的均值表现为通直类型小于弯扭类型。从3个采样地点总苗木的t检验结果可以看出,苗高、分枝高度间的差异达极显著水平 (P< 0.01),地径与针叶长度间的差异不显著 (P> 0.05),且除针叶长度外,其他3个生长量指标的均值均表现为通直类型小于弯扭类型,说明2种茎干类型云南松子代苗木不同个体的4个生长量指标间存在不同程度的变异,且采样地点不同,子代苗木生长的差异性也不同。
综合来看,云南松子代苗木无论是在不同类型间还是在同一类型内各生长指标在不同采样地点间存在着极为丰富的变异性。4个生长量指标中,苗高及分枝高度的变异性是一致的,而苗高和地径间的生长状况不同,可能是由云南松苗木的苗高和地径间生长的异速性造成的,针叶长度作为云南松苗木的一个形态学指标的,其变异相对稳定。
3 结论与讨论
云南松是异花授粉植物,广泛存在着个体之间、林分之间和种源 (产地) 之间的变异。基因型与外界环境条件相互作用共同产生的表型性状变异是物种遗传变异的表现形式,而用形态特征或表型性状来反映遗传多样性是最为传统、简单易行、有效的方法[10-11]。云南松作为西南地区的主要用材树种,其生长和形质性状影响着其木材产品的产量、质量和经济价值,探索2种茎干类型子代苗木生长的变异规律,对于了解云南松的更新、优良种质资源的选育和保存有着重要的意义。
前人曾将树干按通直度划分为不同的等级来分析干形的遗传变异规律[12-13],陈东升等[14]采用树干解析的方法获取干形数据,来分析落叶松种和杂种间干形差异性,实验结果表明干形因子受到一定的遗传控制。周安佩等[15]利用AFLP标记技术对直干、扭曲型云南松进行分析,结果显示2种干形植株间具有明显的遗传分化,也证明了云南松树干扭曲现象受较强的遗传控制。2种不同茎干类型的云南松子代苗木的生长量各指标的分化程度存在一定的差异,但差异不明显,其中,弯扭类型苗高大于通直类型,这与前人的研究结果在树高生长量方面扭转木显著低于正常木不同[6],有可能是云南松茎干弯曲、扭转与其分布地理位置、立地条件和年龄存在密切关系有关[16]。
2种茎干类型在3个采样地点内的苗高、地径、分枝高度及针叶长度4个指标的总平均变异系数存在很大的差异性,变化范围为20.28%~114.27%。在通直、弯扭类型间各性状指标差异性表现也不相同,表现为通直类型各指标的平均变异系数均稍低于弯扭类型,通直、弯扭类型4个指标平均变异系数的变化范围分别为:20.13%~105.90%、20.43%~118.12%,其中,针叶长度的变异系数与前期研究结果相似[17]。苗木各生长量指标在2种类型间的差异性低于类型内各地点间的差异性,但类型间仍存在大的变异,这对云南松苗木来说是有利的,可以使其更适应环境的变化[18]。
对3个采样地点同一类型云南松子代苗木各生长量指标间进行方差分析可知,同一类型内4个生长量指标间的差异性不同,但通直、弯扭类型的各生长量指标间均表现为极显著差异;对3个采样地点的茎干类型云南松子代苗木的生长量指标进行t检验,结果显示,宜良的4个生长量指标在类型间均为不显著差异,新平、禄丰的针叶长度均为不显著差异,苗高与分枝高度表现为极显著或者显著差异,苗高与地径间可能因为异速生长而表现不同[19-21]。说明类型内的变异大于类型间的变异,本研究结果与虞泓[22]和邓丽丽等[23]的研究结果类似,即云南松的变异主要来自于群体内,这些丰富的变异性,为云南松优良种源选育奠定了良好的基础。同时也说明不同茎干类型的云南松在类型、采样地点、家系等水平上均表现出多态性,且其苗期的生长状况与其茎干类型并无明显的关系。前人的研究指出,云南松以对母树性状的遗传有利的天然更新为主[24-25],但受人为干扰,致使目前云南松林中扭曲个体的比例较高[26]。因此,虽然茎干类型对云南松苗木生长的影响不明显,但在生产实践中仍建议在云南松择伐中去劣留优,保留优良干型的植株,使优良干型的云南松作为更新母树,进而为云南松茎干类型的改良、提高云南松的利用价值与经济效益提供依据,为优良的云南松种植资源的保存和利用提供物质基础。
猜你喜欢
南京林业大学学报(自然科学版)(2022年1期)2022-11-29
农业机械学报(2020年3期)2020-04-01
种子(2019年5期)2019-07-02
作文周刊·小学四年级版(2019年8期)2019-04-23
国际木业(2018年3期)2018-09-10
农业环境科学学报(2017年11期)2017-12-07
南方农业·下旬(2017年8期)2017-10-23
江苏农业科学(2016年6期)2016-07-25
天然产物研究与开发(2016年11期)2016-06-15
