
蛙源抗菌肽MagaininⅡ对畜禽常见病原菌抑菌活性与体外稳定性测定
2019-01-03 01:53:42徐欣欣施平伟张海文
饲料工业 2018年20期
■徐欣欣 施平伟 张海文,2*
(1.海南大学热带农林学院,海南海口570228;2.海南大学热带动物繁育与营养学实验室,海南海口570228)
抗生素(Antibiotic)是微生物产生的一种生物活性物质,常常作为药物用于人类和动物的疾病预防、疾病治疗和生长促进[1]。在养殖过程中,兽用抗生素常被添加到动物日粮中,以达到提高动物生长速度和饲喂效率[2],抗生素能够起到抑制或杀灭病原菌的作用。目前在我国养殖业普遍存在滥用抗生素的情况,由此导致的药物残留、耐药性以及环境污染等问题亟待解决,寻找能够替代抗生素的物质十分重要。抗菌肽(Antimicrobial Peptides,AMPs),是由生物体特定基因编码产生的,能够对病原微生物的入侵起到防御作用的一类小分子肽[3]。抗菌肽最早被发现是在1974年,瑞典科学家Boman等在惜古比天蚕体内经过诱导后发现了一种具有抗菌活性的物质[4],并于1981年进一步试验后,将其命名为天蚕素(cecropins)[5],之后科学家便纷纷对抗菌肽进行研究。抗菌肽MagaininⅡ是由Zasloff等在非洲爪蟾(Xenopus laevis)皮肤中发现的[6]。本试验通过研究抗菌肽MagaininⅡ对畜禽常见病原菌的抗菌活性,以此为基础评价其体外不同理化条件下的稳定性,探索其在实际生产中的用途,为MagaininⅡ在畜禽生产上的应用提供参考。
1 材料与方法
1.1 试验材料
蛙源抗菌肽MagaininⅡ(由上海吉尔生化公司合成,纯度达95%以上);阿莫西林(江西省科达动物药业有限公司)、土霉素(购自山西兆益生物有限公司)、庆大霉素(德润通生物科技有限公司)。
表皮葡萄球菌ATCC 12228、金黄色葡萄球菌ATCC 25923、大肠杆菌 ATCC 25922、大肠杆菌O157∶H7、大肠杆菌K12、肠炎沙门氏菌CMCC 50041、铜绿假单胞菌ATCC 27853、鼠伤寒沙门氏菌CMCC 50013和猪霍乱沙门氏菌CMCC 50020(购自中国普通微生物菌种保藏中心)。
1.2 试验方法
1.2.1 菌悬液的准备
将菌株划线接种于MH琼脂平板培养基,37℃培养18~24 h,至长出单菌落;挑取单菌落接种于3 ml新鲜MH液体培养基中,转速250 r/min、37℃恒温震荡培养12 h;转接30 μl上述过夜培养的菌悬液于3 ml新鲜MH液体培养基中,转速250 r/min、37 ℃ 培养2~5 h,分光光度计测定OD600=0.5~0.6;转接10 μl此菌悬液至10 ml新鲜MH液体培养基中,涡旋混匀,此时菌悬液中的细菌数需在5×105~1×106CFU/ml。
1.2.2 抗菌肽稀释液及抗生素稀释液的准备
用无菌96孔平板按规格进行抗菌物质梯度稀释,稀释梯度分别为Magainin Ⅱ:2 560、1 280、640、320、160、80、40、20、10、5、2.5、1.25、0.625 μg/ml共13个浓度;庆大霉素:20、10、5、2.5、1.25、0.625、0.312 5、0.156 2、0.078 μg/ml共9个浓度;阿莫西林:2 560、1 280、640、320、160、80、40、20、10、5、2.5、1.25、0.625、0.312 5、0.156 2、0.078 μg/ml共 16个浓度;土霉素 :20、10、5、2.5、1.25、0.625、0.312 5、0.156 2、0.078 μg/ml共9个浓度。
1.2.3 最小抑菌浓度(MIC)与最小杀菌浓度(MBC)的测定
采用微量肉汤稀释法测定抗菌肽MagaininⅡ以及阿莫西林、土霉素和庆大霉素三种抗生素的MIC和MBC,即按照抗菌物质浓度梯度向无菌96孔板中加入90 μl菌悬液与10 μl抗菌物质并混合,即抗菌物质稀释液再被稀释10倍浓度,阳性对照孔只加入菌悬液不加入抗菌物质,阴性对照空只加入100 μl MH液体培养基,37℃ 静置培养18~24 h。培养结束后,观察孔板各孔底部,没有细菌沉淀产生的抗菌物质对应的最小浓度就是MagaininⅡ以及各抗生素的最小抑菌浓度(MIC)。
从没有细菌生长的孔中吸取10 μl内容物滴至MH琼脂平板培养基,每孔三个重复,37℃培养18~24 h;以是否生长菌落来判定抗菌物质的最小杀菌浓度(MBC)。
1.2.4 蛙源抗菌肽MagaininⅡ与抗生素杀菌速率的比较
以抗菌物质的最小抑菌浓度即1×MIC进行杀菌速率的比较,设置取样时间点为抗菌物质作用后的0、10、30、50、70、90、120 min。
在离心管中依次加入1 ml菌悬液和抗菌物质母液至抗菌物质终浓度为1×MIC,设置空白对照即不加入抗菌物质,置于37℃、转速250 r/min恒温震荡培养;在7个取样时间点,分别取出内容物10 μl,用无菌蒸馏水进行10倍系列稀释,直至稀释到100倍,待混合完全后滴3×5 μl于MH琼脂平板培养基,充分扩散吸收后,培养过夜。
取3个重复总菌落数在15~100的浓度用于菌落计数,计算出原始菌悬液中的细菌浓度,以作用时间点为横坐标,每毫升菌悬液中细菌群落总数(CFU/ml)为纵坐标,做对数图获得时间杀菌速率曲线。
1.2.5 蛙源抗菌肽MagaininⅡ稳定性的测试
1.2.5.1 不同温度对抗菌肽MagaininⅡ抗菌活性的影响
目前,虽然关于我国古代图书收藏与管理等方面的论著颇丰,但笔者认为,古代图书管理与发展过程中,尚存仍需进一步考究与厘清的一些史实和问题。通过查阅相关史料,本文在古代图书馆的产生,古代藏书形式、古代典籍的整理与分类法演变等方面进行了梳理与探讨。
对MagaininⅡ进行梯度浓度稀释,将抗菌肽稀释液置于50、60、70、80、90 ℃水浴加热15 min后取出冷却,测定最小抑菌浓度。
1.2.5.2 胰蛋白酶、胃蛋白酶对抗菌肽MagaininⅡ抗菌活性的影响
对MagaininⅡ进行梯度浓度稀释,分别将胰蛋白酶冻干粉和胃蛋白酶冻干粉按照1∶250的比例和1∶100的比例加入无菌蒸馏水进行溶解。将不同浓度的抗菌肽稀释液分别与胰蛋白酶稀释液和胃蛋白酶稀释液按照1∶1比例加入离心管,置于37℃水浴加热1 h后取出,测定最小抑菌浓度。
1.2.5.3 pH值对抗菌肽抗菌活性的影响
对MagaininⅡ进行梯度浓度稀释,将抗菌肽稀释液与pH值2和pH值8的PBS缓冲液按照1∶1比例混合,测定最小抑菌浓度。
2 结果与分析
2.1 MagaininⅡ与不同抗生素的抑菌活性比较(见表1)

表1 MagaininⅡ以及各抗生素对致病菌的最小抑菌浓度(MIC)和最小杀菌浓度(MBC)
由表1可知,抗菌肽MagaininⅡ以及阿莫西林、庆大霉素与土霉素在相同试验条件下,MagaininⅡ表现出的抗菌活性是最弱的,其对此次试验用革兰氏阳性菌均未发挥抗菌作用,但对几种革兰氏阴性菌表现出一定的抗菌活性,MIC值处于64~256 μg/ml,但对鼠伤寒沙门氏菌CMCC 50013未起到抑菌作用。三种抗生素中,庆大霉素表现出的抗菌活性最强,其对试验菌株都具有较强的抑菌作用,其最小抑菌浓度小于试验所设置的最小浓度,故并未在试验中检测到庆大霉素对这几种试验菌的MIC值,一定程度上体现了庆大霉素较强的抑菌活性;其次是阿莫西林,同样表现出明显抗菌效果,尤其是对大肠杆菌ATCC 25922和大肠杆菌O157∶H7和三种肠炎沙门氏菌,MIC都介于 0.007 8~0.125 μg/ml;土霉素的抗菌活性最弱。此次试验中,MagaininⅡ与抗生素的MBC值都介于其MIC的2~5倍,杀菌活性最强的是庆大霉素。
2.2 MagaininⅡ与不同抗生素的杀菌速率的比较(见图1)

图1 MagaininⅡ与抗生素对金黄色葡萄球菌ATCC 25923(A)和大肠杆菌ATCC 25922(B)的时间杀菌曲线
由图1抗菌肽MagaininⅡ与阿莫西林等几种抗生素对金黄色葡萄球菌ATCC 25923和大肠杆菌ATCC 25922的时间杀菌曲线可知,这几种抗菌物质在最小抑菌浓度下的杀菌速率均不相同,对金黄色葡萄球菌菌落生长抑制作用最为明显是土霉素,其在1×MIC下作用10 min,就在本检测限内检测不到细菌菌落的形成,其次是阿莫西林,需要作用70 min才无细菌菌落形成;而抗菌肽MagaininⅡ表现出不能完全抑制金黄色葡萄球菌菌落的形成,作用50 min后,细菌菌落数便呈现增长趋势并保持增长。对大肠杆菌ATCC 25922菌落生长抑制作用最为明显的是阿莫西林,在最小抑菌浓度下,作用10 min后,就无菌落形成,其次是土霉素,在其作用30 min后,才检测不到细菌菌落的形成,杀菌速度最慢的是抗菌肽MagaininⅡ,在1×MIC下,杀菌速度小于两种抗生素,直到作用90 min才不能检测到细菌菌落的形成。
2.3 MagaininⅡ的体外稳定性测试
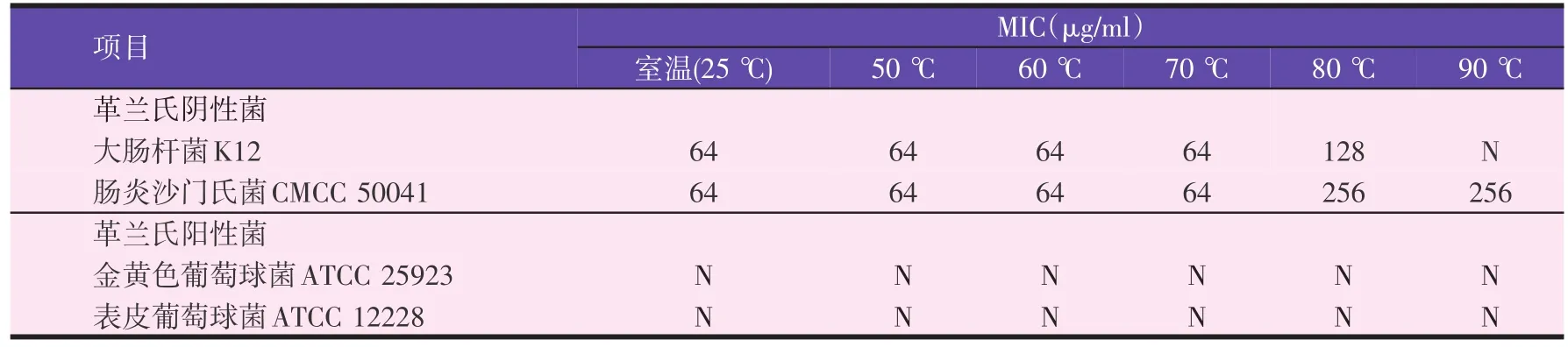
表2 不同温度处理的MagaininⅡ对致病菌的最小抑菌浓度(MIC)
由表3可知,与未处理组相比,经胰蛋白酶和胃蛋白酶处理后的MagaininⅡ不再对肠炎沙门氏菌CMCC 50041和大肠杆菌K12具有抑菌作用,试验中没有检测到其对这两种试验菌的MIC,且处理后的MagaininⅡ依然对革兰氏阳性菌不发挥抑制作用。

表3 胰蛋白酶、胃蛋白酶处理的MagaininⅡ对致病菌的最小抑菌浓度(MIC)

表4 不同pH值处理的MagaininⅡ对致病菌的最小抑菌浓度(MIC)
由表4可知,与未处理组相比,经pH值2处理后的MagaininⅡ仍然能够对大肠杆菌K12和肠炎沙门氏菌CMCC 50041表现出一定程度的抑制作用,但MIC明显增大,经pH值8处理后就检测不到MagaininⅡ对大肠杆菌K12的MIC,但可以检测到对肠炎沙门氏菌CMCC 50041的MIC(128 μg/ml),和未处理组相同的是MagaininⅡ经处理后依然对革兰氏阳性菌不发挥抑菌作用,在试验过程中并未检测到MIC值。
3 讨论
抗菌肽是一种具有抗菌活性的小肽类物质,它在动物和植物体内广泛存在,并在生物进化中发挥了作用,是动物的体内免疫的有效防御武器[7]。抗生素是由微生物在生存环境中产生的能够抑制其它微生物的一类物质,是目前治疗畜禽感染性疾病的常用手段[8]。通过对抗菌肽MagaininⅡ与阿莫西林等几种抗生素的体外抑菌试验结果的比较可以看出,抗生素对试验菌株的抗菌活性好于MagaininⅡ,在此试验中,抑菌效果最好的是庆大霉素,其对革兰氏阳性菌的MIC小于试验设置的最小浓度,MagaininⅡ作为抗菌物质的效果并不明显,目前国内外对于MagaininⅡ抗菌活性的研究,许多是围绕其与其他抗菌肽结合后形成的杂合肽开展,如冯兴军等将牛乳铁蛋白与爪蟾素二者进行连接,并发现两者杂合肽在抑制革兰氏阳性菌与革兰氏阴性菌上都具有明显的作用[9]。
抗菌肽MagaininⅡ与几种抗生素的杀菌速率的试验结果表明,MagaininⅡ在作用金黄色葡萄球菌时,表现为不能抑制该菌的生长,效果不如抗生素;在作用大肠杆菌时,其作用时间也同样长于抗生素,需90 min才可达到有效抑制菌群生长的效果。与阿莫西林和土霉素等抑菌效果较好的抗菌物质进行比较,抗生素在试验中抑制菌群生长所需要的时间则明显少于MagaininⅡ。
MagaininⅡ稳定性的研究结果表明,在经不同温度的处理后,MagaininⅡ在一定的温度范围内,能够保持性质稳定,并未降解;在高温环境中,抗菌活性稍有变弱,但仍然可以发挥抑菌作用。经胰蛋白酶和胃蛋白酶处理的结果表明,与未经处理的MagaininⅡ相比,MagaininⅡ在两种酶处理后,不再对沙门氏菌CMCC 50041和大肠杆菌K12发挥抑制作用,说明在胰蛋白酶和胃蛋白酶的处理下,MagaininⅡ变得不稳定,性质发生改变且抗菌活性变弱。而通过模拟小肠液和胃液的pH值对MagaininⅡ进行不同pH值处理的试验结果表明,MagaininⅡ在酸碱度较高的环境依然能够在一定程度上发挥其抗菌活性,但活性有所减弱,说明MagaininⅡ在不同酸碱度的环境中,能够保持一定的稳定性,发挥对细菌的抑制作用,且在酸性环境较碱性环境性质稳定。
当今抗生素滥用形势严峻,寻找一种能够替代抗生素并且安全无污染的新制剂十分重要。有研究发现,能够在发挥抗菌作用的同时不易产生耐药性的阳离子多肽,具有成为新一代抗菌制剂的潜力[10],抗菌肽由于具有独特的性质,越来越多的研究者纷纷对其进行研究。抗菌肽MagaininⅡ对细菌、病毒等都具有独特的抑制作用,国外许多研究表明,MagaininⅡ不仅对细菌具有抗菌活性,也同样还具有抗原虫的活性,Gwadz等研究表明,MagaininⅡ能够抑制在蚊子宿主体内引起疟疾的寄生虫的卵囊的发育[11],Zasloff则通过对草履虫(Paramecium caudatum)的试验,观察到用MagaininⅡ处理会使得草履虫本身逐渐开始膨胀随后导致破裂,Zasloff等认为在一些生物体内,MagaininⅡ通过扰乱生物膜功能而发挥作用[6]。
国内外有许多实验室研究Magainins的特性,将Magainins与其他抗菌物质进行连接形成杂合肽,进而针对杂合肽的抗菌活性等展开研究,或就Magainins与其他抗菌物质之间的协同效应进行研究,如Glattard等就进行了MagaininⅡ与PGLa混合后的协同增强效应的研究[12]。刘二强等通过研究天蚕素A-马盖宁杂合肽对耐甲氧西林金黄色葡萄球菌的作用,发现这种杂合肽不仅能够抑制该菌DNA、RNA、总蛋白合成的能力,且对细胞能量代谢以及胞内酶活性也能发挥一定的作用[13]。许多研究都表明,将MagaininⅡ与其他抗菌物质连接或混合使用的效果会好于单独使用的效果,在以后的研究中可以充分考虑MagaininⅡ与其他抗菌物质协同效应的研究。
4 小结与展望
MagaininⅡ在此试验中表现出的对革兰氏阳性菌抗菌效果不明显以及在酶处理的情况下性质不稳定等缺点,可以在以后的试验中通过改变其结构或与其他物质联用实现进一步探索,就现阶段而言,抗菌肽的使用还需要大量的评测,目前国内外对于其的研究,主要是研究其与其他抗菌物质进行连接形成的杂合肽的抑菌活性,以及其在免疫调节方面发挥的作用。相信在更深入的系统研究后,关于抗菌肽MagaininⅡ的种种问题与安全性顾虑会得到解决。
猜你喜欢
农家致富顾问·上半月(2021年6期)2021-12-26 17:18:17
现代畜牧科技(2021年9期)2021-10-13 06:38:40
临床医药文献杂志(电子版)(2017年11期)2017-05-17 04:48:36
广东饲料(2016年5期)2016-12-01 03:43:21
国外医药(抗生素分册)(2016年5期)2016-07-12 14:25:37
国外医药(抗生素分册)(2016年5期)2016-07-12 14:25:32
中国现代药物应用(2016年1期)2016-03-04 07:07:42
食品界(2016年4期)2016-02-27 07:37:09
中国洗涤用品工业(2015年11期)2015-02-28 19:03:07
中国药业(2014年24期)2014-05-26 09:00:19
