
水产饲料中酶制剂的研究与应用
2019-01-03 08:40冷向军
饲料工业 2018年22期
■冷向军
(上海海洋大学水产与生命学院,上海201306)
随着全球水产养殖业的快速发展,鱼粉等优质动物蛋白资源的紧缺已成为制约水产养殖业发展的重要因素。开发新的饲料资源,提高现有饲料资源的利用率,是饲料行业所面临的重大问题。近年来,许多动植物蛋白源被开发出来用以降低或取代鱼粉的使用量,比如豆粕、菜粕、棉粕、肉骨粉、血粉等,其中植物蛋白源因其资源量大,价格低而广泛应用于水产饲料中。然而与鱼粉相比,这些植物蛋白源通常具有一种或多种方面的不足,如氨基酸组成不平衡,存在抗营养因子(植酸、非淀粉多糖等)、适口性较差、消化率较低等,限制了植物蛋白源在水产饲料中的大量使用。为提高植物蛋白源的利用率,一些营养策略与技术已被开发并应用于饲料工业中。利用发酵工程和基因工程等生物技术手段开发出的酶制剂,能够提高饲料的利用率,在很大程度上缓解饲料资源紧缺的状况。酶制剂因高效、安全、环保,符合资源节约型社会的要求而受到高度重视。
饲用酶制剂是从微生物、动物、植物中提取的具有酶特性的一类物质,其中微生物来源的酶制剂应用最为广泛。饲用酶制剂的种类很多,按照动物自身能否合成,可分为消化酶和非消化酶。消化酶指动物体内能够合成并分泌到消化道进行消化的酶类,也称内源酶,主要包括淀粉酶、蛋白酶和脂肪酶;非消化酶指动物体自身通常不能合成酶类,也称外源酶,包括植酸酶、纤维素酶、木聚糖酶、葡聚糖酶、果胶酶等。按照酶制剂的类型来分,可分为单一酶制剂和复合酶制剂。根据酶的作用底物,可分为蛋白酶、脂肪酶、碳水化合物酶、植酸酶等。
1 植酸酶
在植物性原料中,磷的主要存在形式是植酸磷,包括植酸及其盐类,通常占总磷的70%左右,表1列出了几种常用植物性原料中的植酸磷含量。
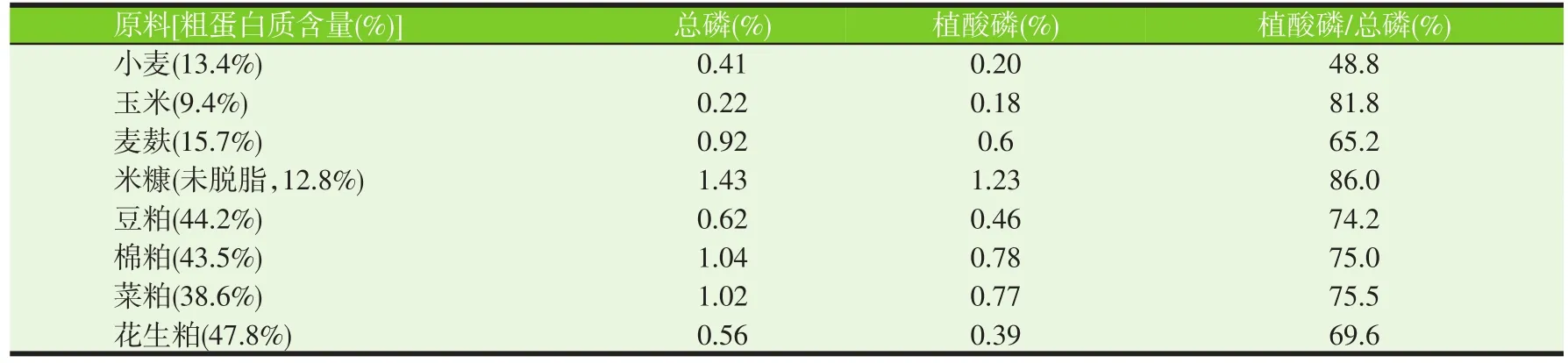
表1 几种植物性原料中的植酸磷含量
对于养殖动物而言,其自身不能分泌植酸酶降解植酸磷,并且植酸磷还会结合其他营养素成分,如矿物质等,降低其利用率。Chowdhury等(2012)发现植酸磷可结合碱性氨基酸如组氨酸、赖氨酸、精氨酸等,从而降低整个氨基酸的利用效率。
植酸酶是催化植酸及植酸盐水解成肌醇与磷酸(或磷酸盐)的一类酶的总称。目前,植酸酶在饲料工业中的应用已有20多年历史,最早是应用于猪、禽动物饲料中,近些年,植酸酶在水产饲料中的使用正在不断扩大。目前在水产饲料中使用的植酸酶主要来自于曲霉(Aspergillus),根据其作用环境,可分为酸性植酸酶和中性植酸酶;根据其形态,有粉末、液体剂型,也有采用包被或微胶囊处理的方式,以增加其稳定性。在使用方式上,可直接在饲料中添加,也有采用后喷涂和原料预处理的方法。
植酸(盐)降解后,可释放出肌醇、无机磷以及与植酸(盐)相结合的营养素,从而提高其利用效率(Cheng等,2002)。Sajjadi等(2004)发现大西洋鲑饲料中添加植酸酶后,缓解了植酸对蛋白质消化带来的负面影响;Liu等(2013)采用原料预处理和后喷涂的方法添加植酸酶,发现当植酸酶的添加量在1 000 U/kg时,草鱼对粗蛋白质、粗脂肪、干物质、灰分、磷和钙的消化率显著提高。类似的结果也见于Zhu等(2015)对黄颡鱼的报道。
饲料中添加植酸酶后,由于提高了磷的利用率,故可减少无机磷的添加,进而节约饲料成本,减少对环境的磷排放。Cheng等(2016)发现,黄颡鱼饲料中50%的磷酸二氢钠可以被植酸酶(1 000 IU/kg)取代,并且由于氮、磷利用率的提高,氮、磷排泄减少了25%和31%。Li等(2018)将罗非鱼饲料中磷酸二氢钙的添加量从1.8%(对照组)降为1.2%,同时补充中性植酸酶0.4 g/kg(2 000 U/g),鱼体增重和饲料系数与对照组没有差异;在磷酸二氢钙含量为1.0%的饲料中添加800 IU/kg中性植酸酶可促进异育银鲫生长,提高干物质和磷的表观消化率,800 IU/kg中性植酸酶在底物充足条件下可代替0.5%磷酸二氢钙的添加(张晓清等,2016)。
目前,大部分商业植酸酶均为酸性植酸酶,其最适pH值为4.0~6.0,这就限制了此类植酸酶在无胃鱼类和虾类饲料中的应用。已有研究表明,在饲料中补充酸性植酸酶对斑节对虾(Biswas等,2007)、日本对虾(Bulbul等,2015)和南亚野鲮(Xavier等,2012)的生长性能并无影响,但补充中性植酸酶却改善了异育银鲫(Liu等,2012)的生长性能和营养物质消化率。故在植酸酶的选择上需根据动物的消化道酸碱性确定。
2 碳水化合物酶
植物细胞和动物细胞的最大差异在于植物细胞具有细胞壁,这是影响植物性原料利用率的主要原因。细胞壁参与维持细胞的形态、增强细胞的机械强度,并与细胞的生理活动有关。细胞壁从结构上可以分为三层:胞间层、初生壁、次生壁。胞间层又称中胶层,为两相邻细胞所共有的一层膜,主要成分为果胶质;初生壁是细胞分裂后,最初由原生质体分泌形成的细胞壁,主要成分为纤维素、半纤维素,并有结构蛋白存在;部分植物细胞在停止生长后,其初生壁内侧继续积累形成次生壁,位于质膜和初生壁之间,主要成分为纤维素,并常有木质存在。可见,构成细胞壁的成分中,绝大部分是多糖,主要是纤维素、半纤维素和果胶类,由葡萄糖、阿拉伯糖、木糖、甘露糖、半乳糖醛酸等聚合而成,另有少量蛋白质。这些细胞壁成分难以消化,在很大程度上阻止了细胞内容物与消化酶的接触,这是植物性原料消化率较低的重要原因。
从动物营养的角度出发,这些细胞壁多糖基本上属于非淀粉多糖(NSP);此外,在细胞内容物中也含有NSP。植物性原料中的NSP含量较高,特别是饼粕类蛋白原料。表2列出了几种常用植物性原料中的NSP含量。根据其溶解性,NSP可分为不溶性NSP(如纤维素)和可溶性NSP(如阿拉伯木聚糖)。近年来,可溶性NSP的抗营养作用日益受到关注。它们与水分子直接作用增加溶液黏度,多糖分子本身也缠绕成网状结构,引起溶液黏度增加,甚至形成凝胶,使得食糜变黏,减少了酶与底物的接触,阻止养分接近肠黏膜表面,从而降低养分消化率。动物消化道缺乏相应的内源酶而难以将其降解。因此,降解NSP,一方面可解除植物细胞壁的屏蔽效应,另一方面可降低黏度,降低其抗营养作用,从而提高植物性原料的消化利用率。
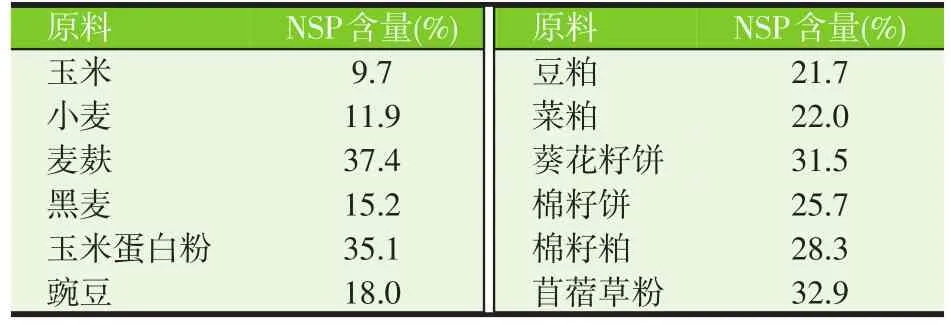
表2 几种植物性原料中的NSP含量
碳水化合物酶是能降解碳水化合物多聚体的一类酶的总称。添加碳水化合物酶可提高生能营养素如淀粉、脂肪的消化率(Adeola,2004);脂肪消化率的提高主要来于碳水化合物酶对NSP的降解,而NSP可以增加胆盐的水解而降低脂肪的消化率(Vahjen等,2007)。此外,碳水化合物酶可以改善摄食高NSP饲料动物的肠道健康。NSP产生的高黏性降低了营养物质的扩散率,导至颗粒物质的积累,使得一些有害生物大量增殖(Vahjen等,1998),通过降低食糜黏度,碳水化合物酶可以消除对肠道微生态平衡产生的不利影响。NSP水解后,产生的寡糖片段,可以为有益微生物利用,通过产生乳酸和有机酸,降低氨的产生(Kiarie等,2007),增加挥发性脂肪酸的浓度(Hübener等,2002),进而改善动物肠道健康和机体健康。
近年来,有关碳水化合物酶在水产营养中的研究开始增多。在草鱼的研究表明,纤维素酶显著提高了生长性能,促进了内源性消化酶的分泌,肠道微生物组成也发生了相应变化(Zhou等,2013);在小麦基础饲料中分别添加不同水平木聚糖酶(0.05%、0.10%、0.15%),显著降低了尼罗罗非鱼的肠道食糜黏度,中、后肠绒毛表面的黏附性颗粒数量减少,颗粒直径减小,中、后肠微绒毛的密度和高度得到不同程度的改善(聂国兴等,2007);在以DDGS部分取代鱼粉的饲料中补充木聚糖酶(20 g/t)和纤维素酶(300 g/t),提高了大菱鲆幼鱼对饲料的利用效果(隋仲敏等,2017)。然而,在以菜粕为主要植物蛋白原料的尼罗罗非鱼饲料中添加纤维素酶,对生长和营养物质利用没有改善(Yigit等,2011);在含10%、20%豆粕的饲料中添加复合酶Superzyme CS(木聚糖酶、淀粉酶、纤维素酶、蛋白酶和β-葡聚糖酶),对虹鳟生长性能没有影响,但降低了粪便黏性和沉降速度(Ogunkoya等,2006)。这些不同的研究结果与所添加酶的种类、数量、饲料组成等因素有关。
有关NSP和碳水化合物的作用机理,在畜禽动物上研究较多,而在水产动物上研究较少。由于生活环境和生理特点的不同,水产动物肠道中的水分含量较高(Refstie等,1999),这使得其黏性较低(Sinha等,2011),可能NSP产生的黏性抗营养作用,在水产动物中不如畜禽动物明显。
目前市场上的主要碳水化合物酶产品是木聚糖酶、葡聚糖酶,占了全球碳水化合物酶市场的80%(Castillo等,2015),其他的一些碳水化合物酶有α-淀粉酶、木聚糖酶、葡聚糖酶、β-甘露聚糖酶、α-半乳糖苷酶、果胶酶、纤维素酶等。
3 蛋白酶
蛋白酶是水解蛋白质产生小肽和氨基酸的一大类酶的总称,蛋白酶的种类甚多,根据其作用环境,有酸性、中性、碱性蛋白酶之分,根据其成分,有单一蛋白酶和复合蛋白酶。目前的蛋白酶产品主要来自于细菌或真菌。饲料中补充蛋白酶,可以弥补动物自身蛋白酶的不足,促进大分子蛋白质降解,提高蛋白质消化利用率。
以植物蛋白原料取代鱼粉后,通常会导致消化道蛋白酶活性的下降。Lin等(2011)以豆粕取代基础饲料(含鱼粉20%)中25%、50%、75%和100%的鱼粉,发现随豆粕用量的增加和鱼粉用量的减少,罗非鱼肠道和肝胰腺蛋白酶的活性极显著下降,而淀粉酶的活性则无显著变化。在凡纳滨对虾日粮中,随大豆浓缩蛋白替代鱼粉比例的增加,肝胰腺蛋白酶活性也表现出下降的规律,脂肪酶、淀粉酶活性与对照组相比没有显著变化(Chen等,2017);类似的报道也见于鲤鱼(冷向军等,2008)。可见,在植物蛋白大量应用和鱼粉用量减少后,补充蛋白酶对于提高植物蛋白的消化利用率具有重要意义。
Drew等(2005)在菜粕-豌豆型饲料中添加250 mg/kg蛋白酶,显著提高了虹鳟对粗蛋白质、脂肪、能量和干物质的消化率;在另一项虹鳟的试验中,在豆粕型饲料中添加228 mg/kg蛋白酶(一种丝氨酸蛋白酶)显著改善了营养物消化率,但在葵花籽粕、菜粕型饲料中补充蛋白酶则没有观察到改善效应(Dalsgaard等,2012);在异育银鲫的研究中,将鱼粉含量从9%降到3%,同时在颗粒饲料中补充蛋白酶后,显著改善了鱼体生长性能,提高了营养物质消化率,但在膨化饲料中补充蛋白酶则对生长和饲料利用没有影响,其原因可能在于膨化加工过程中的高温高压已使大部分蛋白酶失活,再者,饲料膨化后提高了营养物质的利用率,已经能满足鱼体的营养需要,故补充蛋白酶后没有观察到改善效应(Shi等,2016)。在凡纳滨对虾(Li等,2016)、罗非鱼(Li等,2018)上也有补充蛋白酶后改善了鱼体生长性能和饲料利用的报道。然而,在羽扇豆(脱壳)型饲料中补充蛋白酶,对虹鳟生长性能并无改善(Farhangi等,2007)。可见,蛋白酶的作用效果与其底物密切相关。
此外,一些研究表明,在虹鳟(张娟娟等,2012)、罗非鱼(Li等,2018)饲料中补充蛋白酶后,增加了肠绒毛的高度,改善了肠道组织结构。
4 脂肪酶
脂肪酶可将脂肪水解为甘油一酯、甘油二酯和游离脂肪酸,是动物体内的一种重要消化酶。杨新文等(2010)在饲料中添加100、300 mg/kg脂肪酶,显著降低了南方鲇的饲料系数,提高了粗脂肪消化率和蛋白质效率,增重率在数值上也有提高;在瓦氏黄颡鱼饲料中添加300 mg/kg脂肪酶,提高了鱼体增重率9.29%,降低了饲料系数6.35%(谷金皇等,2010);在低蛋白质高脂肪饲料中添加脂肪酶提高了草鱼的生长性能和肠道免疫力(Liu等,2016)。然而在花鲈的研究中,在低蛋白高脂饲料中补充100~800 mg/kg脂肪酶,对生长并无改善,甚至导致了一定程度下降,粗脂肪和部分脂肪酸的表观消化率也下降,这可能是外源脂肪酶添加量过高导致内源酶活性降低,以及脂肪酶水解产生的游离脂肪酸过多而影响了脂肪酸的消化吸收所致(王国霞等,2017)。
总体来看,与植酸酶、蛋白酶、碳水化合物酶相比,脂肪酶在水产上的研究应用比较少,这可能与其稳定性较差、底物不溶于水、生产成本较高等原因有关。此外,除冷水性鱼类和肉食性鱼类外,水产饲料中的脂肪水平并不高,且水产动物对脂肪具有较高的吸收利用率,脂肪酶在水产饲料中的重要性似乎不及其他几类酶制剂。
5 酶制剂使用中的问题与展望
5.1 酶的稳定性与添加方式
酶制剂应用于饲料中,最大的关注点是其稳定性,包括饲料加工中的温度、水分、压力对酶活性的影响,饲料贮存过程中酶活性的损失,酶进入动物消化道后,能否耐受胃酸的影响(有胃鱼类)而到达小肠的作用部位。开发稳定性的酶制剂一直是酶制剂工业努力的方向。可以采用稳定化技术来提高酶的耐热性,如物理包埋、化学修饰等方法;也可采用基因工程等生物技术手段,开发出稳定的酶制剂,或在极端环境中寻求耐高温微生物,产生耐高温的酶。
根据酶制剂的稳定性和饲料生产方式,酶制剂的添加主要有直接添加、制粒后添加和在原料中添加(原料预处理)三种方式。
直接添加:使用粉状饲料时,可以直接添加酶制剂,混合均匀后进行饲喂。如是耐高温的酶,也可以直接加到饲料中再制粒。
制粒后添加:为了避免调质、制粒过程中造成酶活性的损失,可采用制粒后喷酶技术,通常采用液体酶,通过喷雾系统,将酶均匀地喷涂到颗粒饲料表面。
原料中添加(原料预处理):饲料原料在使用之前用酶制剂进行水解,可降解部分抗营养因子,提高饲料的营养价值。如使用纤维素酶、木聚糖酶等碳水化合物酶降解细胞壁和NSP,使用植酸酶降解植酸,使用蛋白酶降解大分子蛋白,这些酶解过程可单独进行,也可复合进行,但前提是这些酶具有相近的水解条件。这些酶解技术,是生产酶解豆粕等原料的理论基础。相比较而言,利用植酸酶对原料进行预处理的方式较为普遍。在大多数的研究中,植酸酶的作用温度和pH值分别为50~55℃和5.0~5.5,也有少量报道为30~37℃温度和微碱性pH值。
5.2 酶的作用环境
酶的水解作用,需要一个适宜的温度、酸碱性和水分环境。大多数商业植酸酶的适宜水解温度是40~60℃,显然水产动物体内无法提供如此高的温度,特别是冷水性鱼类的体温更低。在水产动物消化道内,酶的水解效果难以达到其最佳水平。开发作用温度较低的酶类,以及研究在水产动物生理条件和温度下酶的水解作用,是今后酶制剂研究和应用的一个方向。
鱼类的消化道组成具有很大的多样性,有的有胃,有的无胃;虾类的消化道又具有与鱼类不同的特点,比如虾的胃主要进行机械性消化,其主要的消化吸收部位是肝胰腺。不同的消化道结构特点,使其酸碱性表现不一,因而在选择酶制剂时,需注意酶的作用环境是酸性、碱性或中性,即选择酸性、碱性或中性酶制剂。
5.3 酶的作用底物与基础饲料组成
酶的作用具有高度专一性,只能作用于具有特定化学键的底物。饲料原料结构不同,酶作用的底物不同,因而效果不同。要充分发挥酶制剂的作用,应根据饲料组成而选择相应的酶制剂。比如,在虹鳟的豆粕型饲料中添加蛋白酶显著改善了营养物质的消化率,但在葵花籽粕、菜粕型饲料中补充同样的蛋白酶,则没有观察到改善效应(Dalsgaard等,2012);在选用碳水化合物酶降解NSP时,首先要清楚基础配方中主要的NSP和水溶性NSP是什么,然后针对性的选择木聚糖酶、甘露聚糖酶、葡聚糖酶等。
基础配方的营养水平会对酶的作用效果会产生显著影响。Kumar等(2006)发现,在粗蛋白质含量27%的饲料中补充α-淀粉酶,改善了南亚野鲮的生长和蛋白质利用,但在35%粗蛋白质饲料中补充则无改善效应;在含未糊化淀粉饲料中补充淀粉酶,观察到了对生长的改善效应,但在含糊化淀粉饲料中补充则无改善;其原因可能与糊化淀粉易水解导致葡萄糖吸收过快而产生代谢应激有关。冷向军等(2008)在鱼粉含量10%的鲤鱼饲料中补充蛋白酶显著提高了鱼体增重和饲料利用,但在鱼粉含量15%、20%的饲料中补充蛋白酶则未对鱼体增重和饲料利用产生显著影响。可见,在营养水平较低的基础饲料中补充酶制剂,才能产生较好的改善效果。
5.4 酶的复配
植物性原料中含有多种抗营养因子,单靠一种酶是难以降解的。以植物细胞壁为例,要实现其完全降解,需要纤维素酶、半纤维素酶(包括木聚糖酶、甘露聚糖酶等)、木质素酶、果胶酶四大系列酶类,在细胞内容物暴露出来后,还需要NSP酶、植酸酶、蛋白酶等,显然多种酶的复合,其效果优于单一酶制剂。然而,多种酶复合后,并不是每一种酶都发挥着同等重要的作用。Tahir等(2008)评估了单独或复合添加半纤维素酶、纤维素酶和果胶酶的作用效果,发现半纤维素的水解是整个酶解过程的限速步骤,这意味着纤维素酶和果胶酶在NSP降解方面发挥的作用较少。酶制剂的复配效果与饲料原料组成有关,今后应就酶制剂的单一添加和复合添加的效果进行比较研究。
多种酶制剂的复配,并不意味着要将植物细胞壁和NSP完全降解为单糖,实际上,细胞壁和NSP的部分降解已经达到了破坏抗营养因子的目的,并且产生的寡糖片段具有益生元的功能,可促进肠道有益微生物的生长,改善肠道微生态平衡。
5.5 酶在饲料加工过程中的作用
传统观点认为,酶制剂在进入动物消化道后才开始发挥作用,但在生产中,一些酶制剂在调质、制粒、后熟化后,其活性已大部分消失,但饲养试验的效果却不错,这表明,酶的作用不只局限在动物体内,在饲料加工的过程中就已经开始发挥作用了。Li等(2016)用试验证明了在制粒和熟化过程中,蛋白酶已经发挥了水解蛋白质的作用,该蛋白酶经过制粒和后熟化过程后,活性可保留79.3%和67.5%。这提示我们,在饲料调质过程中,可以通过延长调质时间,适当降低调质温度,从而更充分的发挥酶的水解作用,实际上,这也是原料预处理的一种方式。当然,这种方式的前提条件是所用酶制剂具有较高的耐温性。
总体来看,酶制剂在水产饲料中的应用,已经取得了一定成效。随着稳定性酶制剂和符合水产动物生理特点酶制剂的开发,酶制剂将在水产饲料中得到更多的应用,从而提高饲料利用率,节约饲料资源,促进水产养殖业的可持发展。
猜你喜欢
中国饲料(2022年13期)2022-07-07
中国畜牧杂志(2020年11期)2020-11-18
国外畜牧学(猪与禽)(2020年8期)2020-03-04
生物技术通报(2019年7期)2019-02-21
猪业科学(2018年5期)2018-07-17
中国畜禽种业(2018年1期)2018-01-18
广东饲料(2016年2期)2016-12-01
畜牧兽医科技信息(2015年5期)2015-12-27
浙江农业科学(2012年10期)2012-12-24
养殖与饲料(2012年10期)2012-12-10
