
利用TALEN技术制备HE4(WFDC2)敲除小鼠模型*
2019-01-02 10:43龙华洋李金萍江世文
实验动物科学 2018年5期
龙华洋 孙 丹 李金萍 江世文,3
(1.首都医科大学基础医学院遗传与发育生物学系,北京 100069)(2.第四军医大学唐都医院妇产科生殖医学中心,西安 710038)(3.基础医学系,默索大学医学院,纪念健康医学中心,妇科与妇产科系,萨凡纳,GA,美国,31404)(4.常州大学制药与生命科学学院,常州 213164)
人附睾蛋白 4(Human epididymis protein 4,HE4)最初是在人附睾中被发现,并因此而得名[1]。此后HE4被证明是由人 Wfdc2(WAP four-disulfide core domain protein 2)基因编码的一种分泌蛋白,因此HE4又名为 WFDC2[2]。另有研究报道,除了在人附睾中表达外,HE4也在其他正常组织器官中有表达,包括生殖系统,呼吸系统,并且HE4在气管中表达最高,其余依次唾液腺,肺等组织[3-4]。
近年来,越来越多的研究指出,HE4在一些癌症中高表达,包括卵巢癌[3,5-7],肺癌[8-11],子宫内膜癌[12]。因此认为HE4可以作为检测这些癌症的标志物。在HE4相关癌症的研究中,HE4与卵巢癌的研究最受研究者关注。目前,CA125的含量仍然是检测卵巢癌的金标准[13],Hellstrom等[5]用卵巢癌患者血清中的 HE4水平来检测卵巢癌,其结果与CA125有相同的灵敏性,并且用HE4作为检测指标能提高恶性肿瘤检测的特异性。Ghasem i[14]和Zhao等[15]指出,CA125联合 HE4检测可以提高卵巢癌检测的敏感性和特异性。研究指出,HE4可能是促进卵巢癌细胞[16-17]和子宫内膜癌细胞[18]增殖的一个激活因子。而且,在癌细胞裸鼠异种移植实验中,过表达HE4的肿瘤体积明显大于对照组肿瘤[17]。上述研究提示,HE4高表达与肿瘤增殖有关,但是在正常组织中,HE4缺失的影响以及它正常的生理功能仍然不清楚。
基因敲除小鼠模型是研究动物基因功能的重要工具之一,小鼠的基因组与人类基因组相似度达90%,有99%的基因与人类基因同源[19],并且表达HE4蛋白的人Wfdc2在如小鼠等一些物种间高度保守[3],但目前尚无 WFDC2敲除小鼠模型来研究HE4的正常生理功能。
基因敲除技术的传统方法是通过同源重组获得基因敲除的ES细胞,然后通过嵌合体技术获得基因敲除动物[20],近几年来兴起的基因编辑工具包括锌指核酸酶 ZFN技术[21],转录激活因子样效应物核酸酶TALEN技术[22],成簇规律间隔的短回文重复序列CRISPR以及CRISPR相关蛋白系统CRISPR/Cas9技术[23]。本研究采用TALEN技术敲除C57BL/6 J小鼠Wfdc2基因,首次获得WFDC2敲除小鼠模型,并发现纯合WFDC2敲除小鼠出生后短时间内死亡,为进一步研究HE4的正常生理功能奠定基础。
1 材料与方法
1.1 实验动物
实验动物由北京生命科学研究所转基因动物中心提供。使用许可证号:SYXK(京)2015-002。所有实验小鼠均在SPF级动物房饲养和繁殖。温度控制在 22℃,湿度 70%,自动光周期(12 h明/12暗),自由采食和饮水。
1.2 主要试剂
Golden Gate TALEN assembly合成的质粒(Addgene),10X T4 DNA ligase buffer(NEB),T4 DNA ligase(NEB),限制性内切酶 Bsa I(NEB),Sac I,Esp3I(Fisher)质粒小提试剂盒、普通琼脂糖凝胶DNA回收试剂盒、普通DNA产物纯化试剂盒、T载体试剂盒、mMESSAGE mMACHINE T7 Ultra Kit(Life Technologies)。
1.3 TALENs位点设计
根据 NCBI上小鼠 Wfdc2基因序列,利用TALEN在线设计工具(http://tale-nt.cac.cornell.edu)设计基因敲除位点。TALENs敲除靶序列如下:TALEN-L靶向序列5'-GCA CCA TGC CTG CCT G-3',TALEN-R靶向序列 5'-AGT AGG AGG CCA GCG GC-3'。中间的 spacer序列为 5'-TCG CCT CTG CTT GCT G-3'。
1.4 TALENs载体构建
采用Golden gate法将识别靶序列的RVD模块构建到载体上。然后将TALEN片段通过 Esp3I酶切构建到pCAG-T7-TALEN载体上,使得TALEN表达有 CAG或者 T7启动子启动,其中 CAG启动TALENs在动物细胞中表达,T7启动 TALENs体外转录成mRNA。将测序正确的TALENs表达载体采用无内毒素质粒提取试剂盒提取质粒备用。
1.5 体外转录
Sac I酶切 TALENs无内毒素质粒,电泳回收片段,并进行纯化。以纯化片段为模板,采用mMESSAGE mMACHINE T7 Ultra Kit进行体外转录合成多聚腺苷酸化的mRNA。将体外转录的稳定mRNA用NucAway Spin Columns纯化回收并用显微注射缓冲液稀释到100ng/μL,-80℃保存备用。
1.6 TALENs显微注射及活性验证
将WFDC2的TALEN-L和 TALEN-R mRNA混合在一起后在显微镜(Nikon)下利用显微操作系统(Eppendorf)注射到15枚 C57BL/6 J的受精卵中,培养3.5 d后挑选出存活的囊胚,混合一起用裂解液裂解,以裂解液为模板,以上游引物:5'-TGTGCTTCCTACCCGCCTCCT-3'和下游引物:5'-CCCGCTCCTCTTCATCAGGTCTC-3'进行PCR扩增,并采用T7E1进行酶切鉴定,其中扩增片段大小为413 bp,酶切片段为120 bp+293 bp。验证 TALENs有切割活性后,按照上述方法,获得显微注射受精卵,将其移植到ICR代孕雌鼠输卵管内。
1.7 突变检测及基因型分析
取1周龄小鼠脚趾提取基因组DNA,同时进行编号,按照酶切鉴定方法对F0代小鼠进行基因型鉴定,并将T7E1酶切鉴定为阳性的小鼠的PCR扩增产物连接到T载体上,进行测序和突变基因分析。将Wfdc2基因发生移码突变的F0代小鼠单独建系,最后得到Wfdc2纯合双敲除小鼠。1
.8 WFDC2敲除小鼠表型分析
对WFDC2敲除小鼠的出生情况进行观察分析,记录每一窝的出生胎鼠数量,基因型比例以及存活情况。
2 结果
2.1 TALEN载体构建
小鼠Wfdc2基因有五个已知转录变异体,为了高效敲除WFDC2蛋白表达,本研究选择的敲除位点为 Wfdc2基因的第一个外显子区域,如图 1。TAL-effector识别模块 HD、NG、NI、NN分别识别碱基C、T、A、G,因此本研究中识别 Wfdc2的模块序列分别为5'-NN HD NIHD HD NING NN HD HD NG NN HD HD NG NN-3'和TALEN-R 5'-NINN NN NI NN NN HD HD NINN HD NN NN HD HD NI-3'。采用Golden gate的方法[24]获得了 TALENs体外转录模板,如图2。
2.2 WFDC2敲除小鼠构建及鉴定
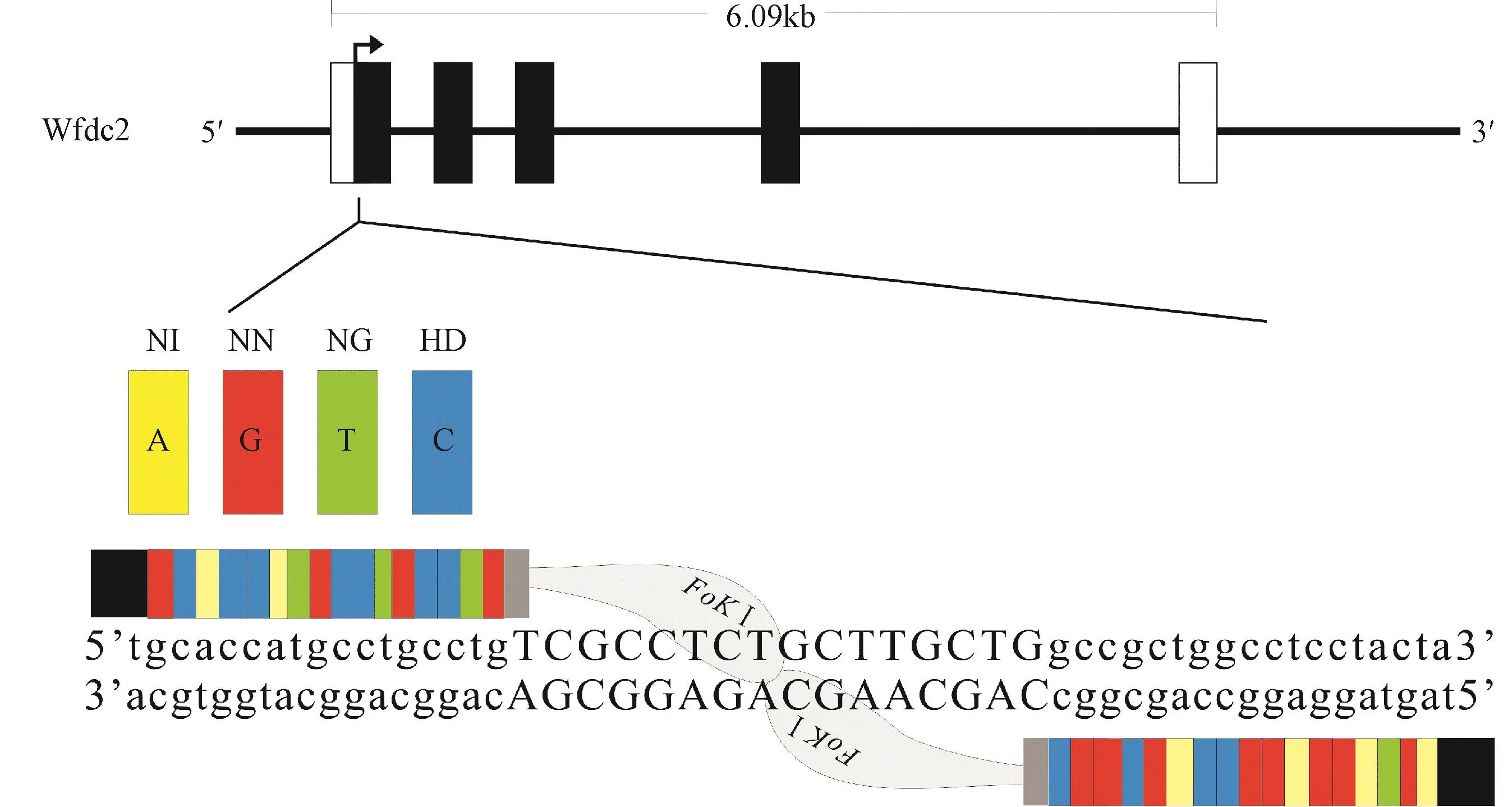
图1 Wfdc2基因敲除TALENs位点Fig.1 The TALENs binding site in the m ouse Wfdc2 gene
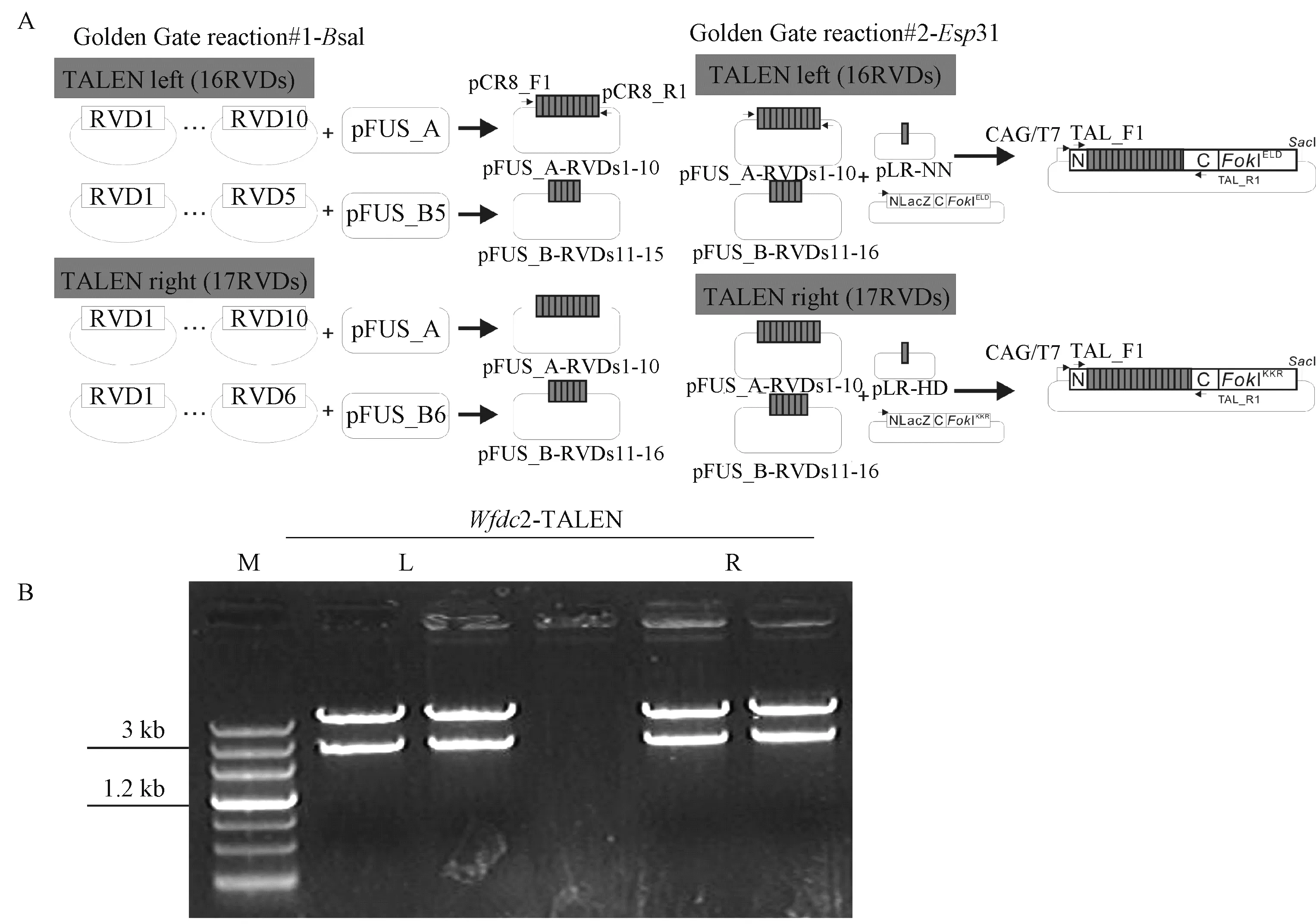
图2 Wfdc2 TALENs识别臂的构建Fig.2 The construction of Wfdc2 TALENs tem p late
体外转录分别得到 TALEN-L和TALEN-R的mRNA。将mRNA分别稀释到100ng/μL后等量混合进行胞质注射,先注射15枚受精卵,进行TALENs活性验证(图3),验证TALENs有切割活性后,共注射140枚受精卵,移植131枚到5只ICR代孕雌鼠输卵管中,共出生24只小鼠。对出生的24只F0代小鼠进行鉴定,鉴定结果显示,有9只敲除阳性小鼠(图4),选取第24号 F0代阳性小鼠继续繁育最终获得敲除17个碱基的Wfdc2纯合双敲除模型。
2.3 WFDC2敲除小鼠的表型分析
得到24号F0代阳性小鼠后,我们将它与野生型小鼠交配,F1代留下敲除17个碱基的杂合WFDC2敲除小鼠(WFDC2+/-),这些杂合小鼠能正常生长,和野生型小鼠生长情况并无明显的差别,WFDC2+/-小鼠自交,能正常怀孕产仔得到F2代小鼠,根据孟德尔第一定律,F2代小鼠中将出现野生型(WFDC2+/+),杂合(WFDC2+/-)及纯合(WFDC2-/-)小鼠,并且 WFDC2+/+∶WFDC2+/-∶WFDC2-/-的比例应为1∶2∶1。但是在 F2代小鼠出生后5 d我们对F2代小鼠编号及基因型鉴定时发现,存活的F2代小鼠中没有发现WFDC2-/-小鼠,只发现 WFDC2+/+和 WFDC2+/-。这一结果说明WFDC2-/-小鼠可能胚胎致死或者出生后死亡。

图4 鉴定Wfdc2基因发生切割的小鼠Fig.4 Restriction digest of PCR p roducts used for identification of m ice carrying-induced m utations of the Wfdc2 gene
为验证 WFDC2-/-小鼠是否胚胎致死,我们对7只胚胎 18.5 d(E18.5)WFDC2+/-孕鼠进行剖腹产,发现E18.5 d孕鼠腹中胎儿中有 WFDC2-/-小鼠,并且未见死胎,其中野生型占总出生数量的29.58%,WFDC2+/-占 46.48%,WFDC2-/-占23.94%(表1),接近孟德尔第一定律1∶2∶1的比例。这一结果提示,WFDC2-/-小鼠可能在出生后死亡,经过仔细观察追踪,我们发现,每窝新生胎儿中,都有一些新生小鼠尸体或残骸,怀疑有的可能被母鼠吃掉。接着,我们对这些尸体或残骸进行基因型鉴定,发现这些死去的胎鼠是WFDC2-/-小鼠。为了防止母鼠将新生 WFDC2-/-小鼠吃掉,我们将刚出生的胎鼠立即与母鼠分开,对不同 WFDC2+/-自交所产生6窝新生小鼠出生情况进行统计,其中50%的新生WFDC2-/-小鼠在5 h之内死亡,所有新生WFDC2-/-小鼠在18 h内死亡,而 WFDC2+/-小鼠和WFDC2+/+小鼠能正常存活下来(图5);在这些胎鼠中,WFDC2+/+占总出生数量的 23%,WFDC2+/-占 52%,WFDC2-/-占 25%(表 2),接近孟德尔第一定律;而 WFDC2+/-自交后代每窝平均产仔数和WFDC2+/+自交后代每窝平均产仔数之间没有显著差异(图 6)。上述结果说明非纯合WFDC2敲除小鼠和野生型小鼠一样具有正常的生长发育和生殖繁育能力,而纯合 WFDC2敲除小鼠能够正常出生,并在出生后死亡。
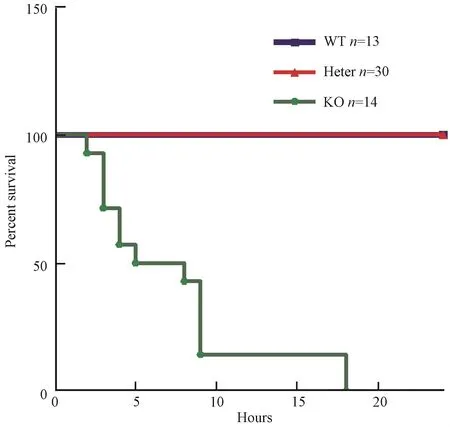
图5 新生小鼠24 h内生存曲线Fig.5 Survival curve for WT、hetero and KO fetus after birth w ithin 24 hours
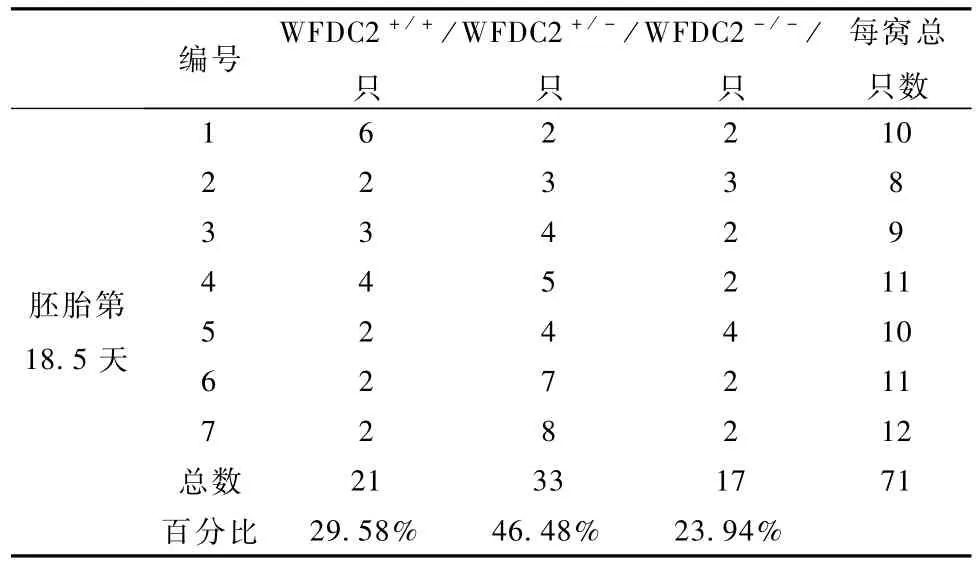
表1 E18.5天胎鼠基因型分布情况Tab le 1 Genotype distribution of em bryos from E18.5
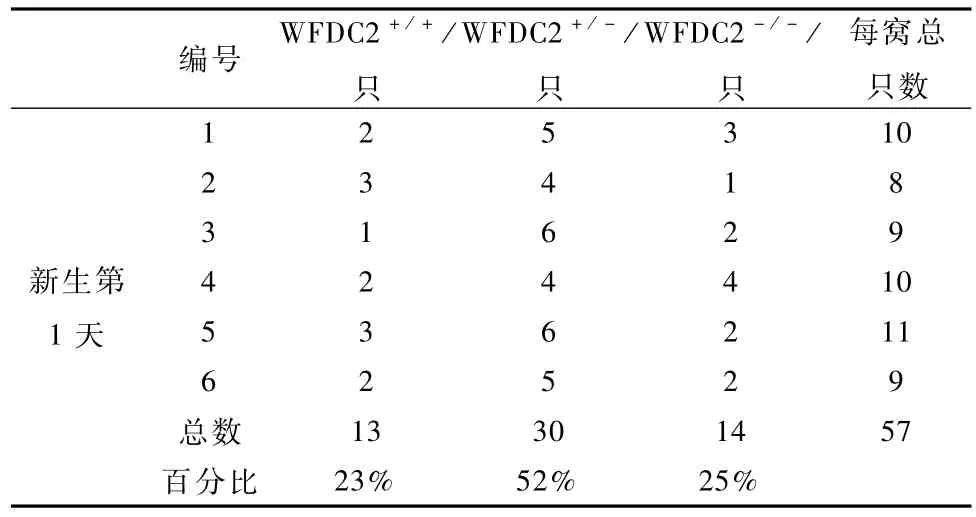
表2 新生小鼠基因型分布Tab le 2 Genotype distribution of neonatal from P0

图6 WFDC2+/-自交后代每窝平均产仔数和WFDC2+/+自交后代每窝平均产仔数Fig.6 Summ ary of litter sizes born from WFDC2+/+and WFDC2+/-
3 讨论
现今基因组编辑技术突飞猛进,在TALEN基因编辑技术之后,又出现了CRISPER-Cas9基因编辑技术,因其操作周期短,效率高而被广泛使用[23,25]。而TALEN也因其相对操作简单,特异性高等特点,仍被广泛应用于动植物等的基因编辑。本研究通过胞质注射TALEN mRNA的方法获得了Wfdc2基因发生移码突变的F0代小鼠,将该种小鼠单独建系,繁育和培养,最后获得纯合的WFDC2敲除模型小鼠。
现今有关WFDC2功能的研究,特别是它在肿瘤发生过程中所起作用的研究越来越受人重视。许多研究都指出卵巢癌,肺癌等组织中HE4有异常高表达,并提出 HE4可以作为这些癌症的检测指示物。但HE4的高表达是促进癌症的发生还是由于癌症的发生而导致HE4的高表达这个问题仍然没有定论。在目前的研究更倾向于HE4高表达促进癌症的发生。Zhu等[16]通过沉默卵巢癌细胞中HE4的表达,发现这些癌细胞的增殖及迁移能力随HE4的减少而降低,指出HE4的表达可能是维持肿瘤细胞增殖、迁移的因子,并且Moore等[17]通过在卵巢癌细胞中高表达HE4,证明HE4促进卵巢肿瘤的发生。但是 Gao等[26]和 Kong等[27]却指出 HE4在肿瘤中起着抑制癌细胞增殖和迁移的作用。因此现阶段关于HE4是否促进肿瘤的发生的结论还需要更多的研究来证实。WFDC2除了在肿瘤组织中有高表达外,它在正常组织中也有表达。在人体正常组织中表达量从高到低的器官依次是气管,唾液腺,肺;而在小鼠正常组织中表达量从高到低的器官依次是肺,肾等[4]器官,这些研究结果提示HE4可能参与调控体内相关正常生理活动。
本研究发现WFDC2纯合敲除小鼠出生后死亡,而非纯合WFDC2敲除小鼠能正常生长和繁育。可能的原因是:1).纯合WFDC2敲除小鼠在孕鼠母体内能够获得来自母体血液的WFDC2,而出生后由于缺乏母体和自身合成的WFDC2而死亡。而非纯合小鼠由于未全部失去自身合成WFDC2的能力而能在出生后正常存活并生长发育。有趣的是,WFDC2在一些物种中高度保守[3],成熟WFDC2蛋白分子量在13kD,在被糖基化修饰后为27kD左右,至于27kD的WFDC2是否能透过胎盘屏障而到达胎儿体内还有待实验验证。2).WFDC2表达可能具有时空性,在胚胎发育期或胚胎发育早期,WFDC2不参与相关生理活动,在小鼠出生后 WFDC2参与并维持正常生理活动,而纯合敲除小鼠由于缺乏WFDC2表达而不能实现相关生理功能正常进行,最终导致死亡。本实验首次通过实验证明WFDC2参与调控小鼠正常生理活动,提示 WFDC2可能在维持胎儿正常生理功能中起着重要的作用,同时,通过研究HE4正常生理功能的作用机制,能更好的帮助我们理解它们在癌症发生中所起的作用。因此,本研究获得的WFDC2敲除小鼠是研究WFDC2生理功能的良好模型。
猜你喜欢
昆明医科大学学报(2022年1期)2022-02-28
昆明医科大学学报(2020年12期)2021-01-26
猪业科学(2018年4期)2018-05-19
安徽医科大学学报(2016年12期)2017-01-15
广东饲料(2016年4期)2016-12-01
广东饲料(2016年8期)2016-02-27
肿瘤预防与治疗(2015年2期)2015-09-26
医学研究杂志(2015年4期)2015-06-10
现代检验医学杂志(2015年6期)2015-02-06
作物研究(2014年6期)2014-03-01
