
放牧与舍饲育肥对绒山羊成年羊与羔羊瘤胃内几种微生物数量的影响
2019-01-02 12:04孙国平闫素梅
饲料工业 2018年16期
■张 娟 孙国平 韩 帅 闫素梅
(内蒙古农业大学动物科学学院,内蒙古呼和浩特010018)
阿尔巴斯白绒山羊是世界著名的绒肉兼用型品种,所产羊绒以细、长、柔软著称,素有“纤维宝石”之美誉,一直以来是饲养绒山羊主要的经济增长点[1],然而,近几年羊绒价格的下跌导致了山羊育肥成为了新的经济增长点。在绒山羊的饲养过程中,大量被淘汰的母羊和羔羊被用来育肥做肉用。相关文献报道每年留作种用的羔羊仅30%,且每年淘汰30%左右成年羊作育肥用,成为羊肉的主要来源[2-3]。现如今,生态环境极具脆弱与敏感,加上人为干扰,使生态环境问题越来越严峻,限制了放牧养殖,这就使得短期舍饲育肥成为了绒山羊育肥方式的新的关注点[4]。瘤胃发酵功能直接影响反刍动物的生产效率与产品品质,育肥方式的改变会对日粮精粗比产生影响,且通过瘤胃微生物的变化对瘤胃发酵功能的影响较大,从而对育肥效果及肉品品质等产生影响,但究竟如何影响尚不清楚。瘤胃是反刍动物特有的消化器官,反刍动物可通过瘤胃中细菌、原虫和真菌发酵分解饲料进而获得营养物质[5]。杨蕾等[6]指出瘤胃中微生物与宿主之间存在互惠共生的关系,瘤胃中所含的26类、7 000多种菌之间的相互作用,影响蛋白质、脂肪等沉积速率以及纤维素的降解速率,从而改善反刍动物消化功能,影响其肉品品质。郭秀兰[7]发现脂肪沉积与硬壁菌门呈正线性关系,与拟杆菌门呈负线性关系,这说明瘤胃内菌群的变化会影响脂肪沉积,进而影响肉品质及其风味。课题组前期就自然放牧与舍饲对绒山羊生产性能及肉品品质的影响做了大量研究,结果表明舍饲可提高绒山羊饲料转化效率、生产性能和屠宰性能,对理化特性并未产生不利影响,但影响羊肉的氨基酸与脂肪酸组成[1,3,8-9],其机理尚不清楚。尹永志等[10]也提出羔羊瘤胃发育并不完善,而瘤胃微生物数量和结构的变化影响机体生理平衡,所以研究羔羊瘤胃内菌群变化对预测其健康成长具有重要意义。本试验旨在探究不同饲养方式对阿尔巴斯白绒山羊成年羊与羔羊瘤胃菌群的影响,为科学的制定阿尔巴斯白绒山羊成年羊与羔羊育肥方案,深入探讨饲养方式对肉品品质的影响机制提供理论基础。
1 材料与方法
1.1 试验动物与试验设计
试验在内蒙古阿尔巴斯白绒山羊种羊场进行。试验分2部分,均采用单因子完全试验设计。试验1将60只年龄、体重相近的淘汰成年母羊随机分为2组,每组30只。一组为对照组,在天然草场进行自然放牧育肥;一组为试验组,完全舍饲育肥,日粮采用全混合饲喂,自由采食,育肥期60 d,1~30 d为育肥前期,31~60 d为育肥后期。试验2从内蒙古白绒山羊种羊场的4月龄断奶羔羊群中选择日龄相近[(130±10)d]的去势公羔60只分为2组,每组30只。一组为对照组,进行放牧补饲育肥,一组为试验组,进行舍饲育肥。放牧补饲育肥的羔羊按照原厂方式,每天在天然草场上放牧并每只补饲玉米(风干基础)300 g,舍饲育肥羔羊按照试验设计的全混合日粮饲喂,各组羔羊均自由饮水。育肥时间为3个月,分育肥前期(1~30 d)、中期(31~60 d)和后期(61~90 d)三期。
1.2 日粮组成
对照组成年羊在天然草场进行自然育肥,采食牧草的主要营养物质含量见表1;试验组的日粮组成及营养水平见表2,日粮干物质基础的精粗比例在育肥前期为40∶60,育肥后期为45∶55。
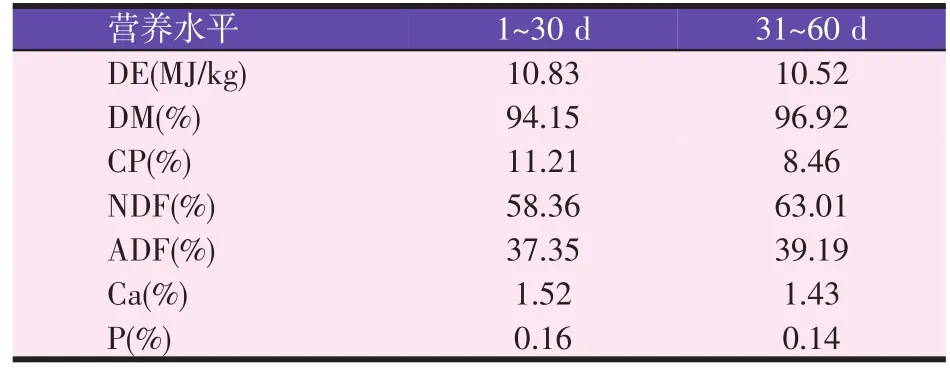
表1 自然放牧成年母羊采食牧草营养水平(风干基础)

表2 舍饲育肥母羊日粮组成和营养水平(风干基础)
对照组羔羊在天然草场进行自然放牧补饲所采食牧草的主要营养物质含量见表3;试验组的日粮组成及营养水平见表4,日粮干物质基础的精粗比例育肥前期为40∶60,育肥中期和育肥后期为50∶50。
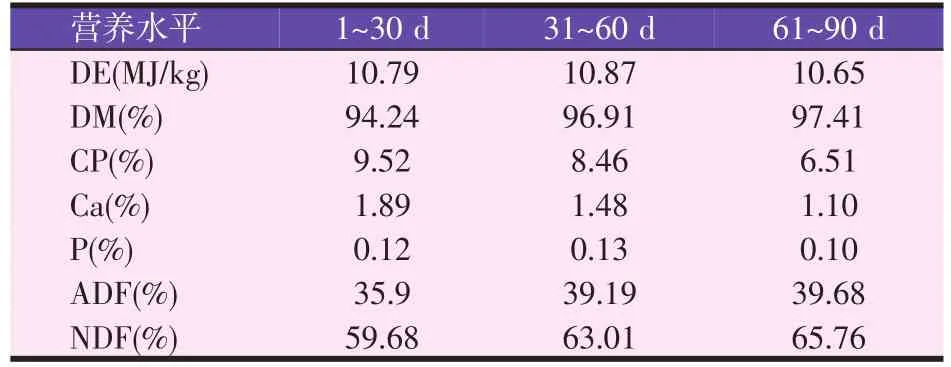
表3 放牧补饲育肥羔羊采食牧草营养水平(风干基础)
1.3 测试指标与方法
在试验结束时,分别从对照组和试验组中选择6只羊、在禁食24 h、禁水2 h后屠宰,采集瘤胃液后立即混匀,将采集的瘤胃液装入到冻存管内,-80℃保存。
1.3.1 瘤胃中总DNA的提取
采用珠磨砚法从处理好的瘤胃液中提取瘤胃液微生物总DNA。DNA的纯度、浓度与玻璃珠的用量和振荡破碎细胞及蛋白质的去除过程密切相关。加入酚∶氯仿∶异戊醇(25∶24∶1)处理5 min,使蛋白质彻底变性并将其完全去除,从而提高DNA产物的纯度。加入氯仿∶异戊醇(24∶1)抽提时,静置后离心是使液面更加清晰,更好的吸取上清,以免蛋白污染。
1.3.2 引物设计、PCR反应体系及反应参数
引物设计及退火温度详见表5。
PCR反应体系为25 μl:模板(瘤胃液DNA)1 μl,目标基因上游、下游引物各1 μl,目标基因下游引物1 μl,DNA连接酶Mix 12.5 μl,ddH2O(无酶水)9.5 μl。PCR反应参数:94℃预变性5 min,95℃变性30 s,退火40 s,72 ℃延伸1 min,40个循环,72 ℃延伸8 min。目标菌株PCR扩增产物经1.5%琼脂糖电泳检测后,均与预期扩增长度一致,可用于胶回收。回收方法参照康为胶回收试剂盒(Gel Extraction Kit,康为世纪生物科技有限公司)的方法进行,产品号为Cat.No.CW0525。

表4 舍饲育肥羔羊日粮组成及营养水平(风干基础)

表5 PCR扩增引物
1.3.3 PCR扩增产物的克隆
按DNA Ligatiog Kit试剂盒说明,将PCR产物与pGM-T载体连接。连接体系为目的PCR片段7 μl、pGM-T 载体 1 μl、0×T4DNA Ligation Buffer 1 μl、T4DNA Ligase 1 μl,共 10 μl。克隆后需要与载体进行连接,为保证连接质量,选择轻弹混匀后16℃过夜。将10 μl连接产物加入到冰浴5 min融化好的100 μl TOP 10感受态细胞中,轻弹混匀后冰浴30 min,42℃水浴90 s后立即冰浴2~3 min,其间不要摇动离心管。向离心管中加入250~500 μl预热的37℃的液体LB培养基(不含抗生素),150 r/min、37℃振荡培养45 min。将离心管中的菌液混匀,6 000 r/min离心5 min。弃去400 μl上清液,抽打混匀剩余菌液取100 μl涂布到含有IPTG、X-gal和含有相应抗生素的LB固体培养基上,无菌玻璃棒均匀的涂开后待平板表面干燥将平板倒置,37℃培养12~16 h。
1.3.4 阳性克隆菌蓝白斑筛选及PCR鉴定
运用蓝白斑筛选法筛选出阳性克隆菌,在LB固体培养基上随机挑选白色菌落,将得到的白色菌落接种于1~5 ml LB培养基(含终浓度为50~100 μg/ml的氨苄青霉素)中,37℃摇床培养过夜。将过夜培养的菌液做普通PCR鉴定,体系及模板量同上。选取经PCR鉴定后的阳性克隆菌液,由中美泰和公司进行测序。测序结果在NCBI的Blast进行比对。
1.3.5 阳性克隆质粒的小量提取及标线制备
对比对成功的阳性克隆菌进行质粒的小量提取,按照质粒小提试剂盒[TIANprep Mini Plasmid Kit,天根生化科技(北京)有限公司]说明步骤进行操作。用酶标仪测定质粒的浓度,再根据公式计算阳性克隆质粒的拷贝数:DNA(拷贝数)=DNA浓度(ng/μl)×6.02×1 023(拷贝/mol)/DNA长度(dp)×660 (g/mol.dp)。然后对已知拷贝数的质粒标准品做至少5个10倍梯度稀释,每个浓度做三个重复,同时用无酶水做对照,用荧光定量PCR仪(BIO-RAD-IQ5TM)按如下体系操作:模板(待测样DNA)1 μl,荧光定量用SYBR Green Ⅱ Mix 12.5 μl、目标基因上游引物、下游引物(10 μmol/l)各1 μl,ddH2O为9.5 μl制备标线。
1.3.6 实时荧光定量PCR分析
所测样本用与上述相同体系及方法进行检测。
1.3.7 瘤胃液的原虫数量
采集的瘤胃液立即用两层纱布过滤,取1 ml该滤液用4 ml MFS染液将原虫固定,静置30 min,常温放置以备用。显微镜计数。
1.4 数据统计
试验数据运用SAS 9.0软件进行t检验,当P≤0.05时,表示组间差异显著,当0.05<P≤0.10时,表示组间差异趋于显著,而P>0.10表明组间差异不显著。
2 结果与分析
2.1 成年羊瘤胃液中的几种微生物数量

表6 不同饲养方式对成年羊瘤胃液中总细菌、总产甲烷菌及三种纤维降解菌数量的影响(copy/ml)

表7 饲养方式对成年羊瘤胃液中反刍月型单胞菌、牛链球菌、嗜淀粉瘤胃杆菌和脂解厌氧弧杆菌数量的影响(copy/ml)
如表6与表7所示,与对照组相比,试验组成年羊瘤胃液中的总产甲烷菌与反刍月型单胞菌趋于显著增加,(P=0.06,P=0.09),牛链球菌趋于显著降低(P=0.07),原虫、产琥珀酸丝状杆菌与脂解厌氧弧杆菌数量显著增加(P=0.01,P=0.05,P=0.05)。成年羊试验组和对照组的瘤胃液总细菌、白色瘤胃球菌、黄色瘤胃球菌及嗜淀粉瘤胃杆菌间差异均不显著(P>0.10),但在数值上试验组高于对照组。

表8 成年羊瘤胃液中的7种菌占总菌的百分比(%)
由表8列出了瘤胃液中的7种菌占总菌的比例。结果得出,试验组的反刍月型单胞菌比例最高,显著地高于对照组(P=0.03),纤维类降解菌中的产琥珀酸丝状杆菌比例次之,显著高于对照组(P=0.03),脂解厌氧弧杆菌占比最低,但显著高于对照组。对照组以嗜淀粉瘤胃杆菌比例最高,反刍月型单胞菌比例次之,脂解厌氧弧杆菌最低。
2.2 羔羊瘤胃液中的几种微生物数量

表9 不同饲养方式对羔羊瘤胃液中总细菌、总产甲烷菌及三种纤维降解菌数量的影响(copy/ml)

表10 饲养方式对羔羊瘤胃液中反刍月型单胞菌、牛链球菌、嗜淀粉瘤胃杆菌和脂解厌氧弧杆菌数量的影响(copy/ml)
如表9、表10所示,与对照组相比,试验组的羔羊瘤胃液总细菌数量显著增加(P=0.003),总产甲烷菌及原虫数量差异趋于显著增加(P=0.08,P=0.09);反刍月型单胞菌、白色瘤胃球菌与黄色瘤胃球菌数量呈上升趋势,其他各菌虽然数值上略有降低,但均无显著差异(P>0.10)。

表11 羔羊瘤胃中7种菌占总菌的百分比(%)
如表11所示,羔羊瘤胃液中7种菌占总菌的比例在两组间均无显著差异(P>0.05),但对照组均略高于试验组。对照组与试验组瘤胃液细菌均以嗜淀粉瘤胃杆菌占比最高,产琥珀丝状杆菌、白色瘤胃球菌次之,脂解厌氧弧杆菌最低。
3 讨论
反刍动物与瘤胃微生物之间建立共生关系,动物通过这些微生物为饲料发酵提供营养和最佳环境条件,进而为宿主动物提供营养物质。如产琥珀酸丝状杆菌是严格厌氧型革兰氏阴性菌,具有较强的纤维素降解能力。可将饲料中纤维物质发酵产生乙酸和琥珀酸。原虫能够将颗粒性淀粉和葡萄糖等可溶性糖等碳水化合物转变成支链淀粉并贮藏于其体内,避免了其他细菌对淀粉和可溶性糖类的快速发酵,以保持瘤胃内pH值的稳定[16]。脂解厌氧弧杆菌可能是目前唯一的脂肪降解菌[17]。是以脂解厌氧弧菌受日粮结构影响较为明显。周航[18]对延边黄牛的研究中发现日粮精粗比对瘤胃微生物群落结构有很大影响。有相关研究表明,羔羊出生后2~4 d可在瘤胃中发现纤维分解菌,说明羔羊在采食固体饲料之前瘤胃中就已经建立了纤维降解菌区系[19]。但建立后的细菌种群或整个微生物区系受日粮影响显著,而Yáñezrui等[20]也通过比较饲喂精料加粗料与仅饲喂粗料对断奶前羔羊瘤胃细菌总数、产琥珀酸丝状杆菌和产甲烷菌数量的影响,说明了幼畜瘤胃微生物区系结构和菌群组成与饲粮类型相关。张海波等[21]及Zhang等[22]均表示随着精补料能量增加,瘤胃原虫数量增多,并提出精补料能量增加可能为原虫的繁殖提供了充足的能量,有利于原虫的繁殖。王梦芝等[23]也表示日粮结构(精粗比)影响瘤胃细菌和原虫数量,本试验中无论成年羊与羔羊,试验组原虫数量均显著或趋于显著的高于对照组,证明饲喂精料后,对原虫数量有影响且有促进作用。课题组前期试验表明完全舍饲显著提高了成年羊瘤胃内乙酸、丙酸水平[24],本试验中成年羊舍饲组产琥珀酸丝状杆菌数量相比放牧组也显著增加,说明舍饲提高了瘤胃内该菌的数量,进而使更多的纤维物质被降解生成乙酸。有研究报道,饲喂高精料日粮可显著增加反刍月型单胞菌的数量,进而提高了丙酸含量,这说明丙酸水平的变化可能与反刍月型单胞菌的数量有关[25-26]。本试验研究结果表明,与对照组相比,舍饲组成年羊瘤胃内反刍月型单胞菌显著增加,这与上述报道一致。这可能是由于舍饲组的日粮营养水平高于对照组,进而增加了成年羊反刍月型单胞菌的数量。而对于羔羊来说,其瘤胃内反刍月型单胞菌变化并不显著,这可能是羔羊瘤胃微生物区系尚不完善的原因。反刍月型单胞菌既是乳酸利用菌[15],也是丙酸生成菌[27],据此可推断该菌可能利用乳酸转化为丙酸,提高瘤胃内VFA的含量,这与本试验结果也相吻合,VFA浓度过高,会导致pH降低,影响瘤胃发酵情况,但本试验同期结果表明pH值在适宜范围内,说明VFA浓度虽然升高了,但并未对瘤胃发酵功能产生影响。
牛链球菌在牛瘤胃中存在广泛,是瘤胃中主要的产乳酸菌和淀粉降解菌[28],该菌可以产生活性较高的脲酶,在瘤胃非蛋白氮的分解中起重要作用。在赵培厅等采用逐渐提高饲粮精料比例的方式诱导奶山羊发生SARA过程中,发现牛链球菌的数量并未随精料比例的改变出现较大变化,且占总菌比例较小[29],这与本试验结果一致。
产甲烷菌利用真菌、原虫发酵过程中产出的H2将CO2还原成CH4,释放到空气中[20];因此,产甲烷菌主要是在瘤胃中起到代谢H2的作用。羔羊在传统模式饲养过程中其瘤胃中产甲烷菌和纤维降解菌很早就被建立起来[19],且有研究也证明产甲烷菌可以影响产氢细菌和原虫群落的数量[30],而产甲烷菌的数量会受日粮碳水化合物的影响。这一点在Johnson等[31]的研究中得到证实,研究指出,给动物饲喂可溶性糖比饲喂淀粉时瘤胃产甲烷菌数量高。本试验结果显示,成年羊和羔羊舍饲育肥条件下相较于放牧或放牧补饲条件下产甲烷菌数量均增加,且差异趋于显著,这是由于试验期在秋季,对照组进行放牧育肥时饲草中粗纤维含量较高,且木质化程度也偏高,牧草适口性差,导致动物采食量下降,瘤胃中不利于产甲烷菌的生长。而完全舍饲组的精料比例较高,可溶性碳水化合物含量高,营养均衡,利于产甲烷菌的生长繁殖,故舍饲组母羊与羔羊瘤胃内产甲烷菌数比对照组多。且羔羊产甲烷菌和纤维降解菌在10日龄即可达到与成年羊类似的水平[20],这也可以解释其菌群变化规律与成年羊相似这一现象。赵玉华[32]提出通过减少原虫数量以减少产甲烷菌附着的载体,可有效减少产甲烷菌,所以原虫数量与产甲烷菌数量变化趋势相一致。原虫数量的增加可能也是导致产甲烷菌数量上升的原因之一。
除产琥珀酸丝状杆菌外,黄色瘤胃球菌和白色瘤胃球菌能将纤维降解后产生大量的纤维素酶和半纤维素酶,其中主要为木聚糖酶。杨宏波等[33]提出黄色瘤胃球菌与白色瘤胃球菌之间有相互抑制作用,可能是白色瘤胃球菌产生的细菌素抑制了黄色瘤胃球菌的生长,且证明这两种菌的变化与日粮结构密切相关。本试验中也发现白色瘤胃球菌数量始终高于黄色瘤胃球菌,这与冯仰廉试验结果也相一致[17]。但本试验中,不同的饲养方式对羔羊与母羊瘤胃中白色瘤胃球菌与黄色瘤胃球菌并无显著影响。马慧忠[34]在对绒山羊进行不同精粗比日粮对其瘤胃发酵影响的研究中也发现这两个球菌的拷贝数发生变化,但组间差异并不显著。日粮精粗比的改变对白色瘤胃球菌和黄色瘤胃球菌的相对表达量也没有影响,这与本试验结果也相类似。瘤胃主要纤维分解菌产琥珀酸丝状杆菌、黄色瘤胃球菌和白色瘤胃球菌只占总瘤胃细菌群落的10%以下,这与本试验结果基本一致[35]。成年羊嗜淀粉瘤胃杆菌变化差异虽不显著,但是舍饲组在数值上高于自然放牧组。这可能是由于嗜淀粉瘤胃杆菌具有降解淀粉的功能,所以有增加的趋势。但精料水平并不足以引起牛链球菌和嗜淀粉瘤胃杆菌的剧烈变化。
本试验结果显示,舍饲条件与放牧条件下羔羊瘤胃中总菌数量差异显著,而成年羊则无显著差异,说明年龄对总菌数有影响,羔羊发育过程中瘤胃菌群可能还不完善,瘤胃内环境还不太稳定。成年羊试验中,除原虫数量、产琥珀酸丝状杆菌数量和产甲烷菌数量在两种饲喂条件下数量差异显著或趋于显著,其他各菌差异均不显著,这说明舍饲在成年羊上效果更为显著。综上所述,结合本试验研究结果表明不同育肥方式下,日粮组成不同会对成年羊及羔羊瘤胃细菌数量及菌群变化产生不同程度影响,且从总体来看,放牧补饲增加了精料的饲喂但结合课题组同期其他方面研究,舍饲育肥较放牧促进了瘤胃微生物的生长,说明舍饲育肥方式可促进绒山羊育肥效果。这可能是舍饲育肥或补饲精料使绒山羊营养摄入更加平衡,且为菌群生长提供了必须能源,利于微生物的生长,从而更好的利用日粮中的营养物质,促进育肥效果。
4 小结
(1)舍饲育肥方式相较于放牧育肥会引起成年羊瘤胃中原虫、产甲烷菌、反刍月型单胞菌、脂解厌氧弧杆菌数量显著或趋于显著升高,三种主要纤维降解菌中产琥珀酸丝状杆菌数量显著升高,而白色瘤胃球菌与黄色瘤胃球菌数量差异不显著。
(2)相较于放牧补饲育肥组,羔羊在舍饲育肥组中总细菌量、原虫数量与产甲烷菌数量显著或趋于显著增加,而其他菌两组间差异并不显著。
猜你喜欢
中国感染控制杂志(2022年11期)2022-11-24
文萃报·周二版(2022年25期)2022-06-23
现代畜牧科技(2021年4期)2021-07-21
新农业(2020年18期)2021-01-07
智富时代(2019年5期)2019-07-05
智富时代(2019年5期)2019-07-05
中国兽医杂志(2019年2期)2019-06-25
中成药(2018年10期)2018-10-26
黑龙江八一农垦大学学报(2017年5期)2017-10-31
湖南农业科学(2017年7期)2017-03-04
