
BRAF在10-姜酚抗黑色素瘤中作用的分子模拟及实验研究*
2018-12-27 07:14张竞之江小梨张双伟
中国病理生理杂志 2018年12期
张竞之, 邓 波, 江小梨, 张双伟, 蔡 敏, 刘 彬
(广州医科大学附属第二医院 1中医科, 2心血管疾病研究所, 广东 广州 510260; 3香港浸会大学深圳研究院, 广东 深圳 518057)
黑色素瘤是最常见的皮肤肿瘤,其发病率和致死率不断升高,由于黑色素瘤的高转移性及高耐药性,被认为是致死率最高的肿瘤之一。据统计,在发达国家黑色素瘤是男性最常见的3种肿瘤之一,占总肿瘤病例的8%[1]。我国黑色素瘤发病率较低,然而近年来由于饮食结构变化及臭氧层破坏诸多因素的影响,我国黑色素瘤发病率发展迅速,据统计2000年我国黑色素瘤发病率为0.2/10万,到2005~2007年其发病率达1/10万[2-3]。目前针对黑色素瘤治疗的药物效果有限,探索新型有效的抗黑色素瘤药物的研发具有重要意义。
生姜是姜科植物姜的根茎,具有解表散寒、温中止呕和化痰止咳等功效。姜酚是生姜的主要活性成分,包括6-姜酚(6-gingerol,6-G)、8-姜酚(8-gingerol,8-G)和10-姜酚(10-gingerol,10-G)[4]。研究表明姜酚类活性成分具有抗氧化、止呕、抗肿瘤、抗炎和免疫调节等作用[5]。姜酚类成分的抗肿瘤作用及其机制一直受到重视[4]。既往研究发现6-G和8-G均可显著的抑制黑色素瘤细胞的增殖[6-7],然而尚未见10-G对于黑色素瘤作用的报道。研究发现10-G具有比6-G和8-G更好的抗肿瘤活性,例如10-G 比6-G能更加显著地抑制白血病细胞的增殖,并且其诱导乳腺癌细胞凋亡的效果比6-G和8-G更加明显[8]。这些提示10-G的抗黑色素瘤作用及机制具有一定研究价值。
BRAF是调控细胞增殖的关键蛋白[9]。研究发现大约50%的黑色素瘤患者出现了BRAF基因突变,其中超过90%为第600位谷氨酸代替缬氨酸的V600E型单核苷酸突变[10]。发生V600E突变后BRAF持续激活,通过不同的机制促进黑素瘤的发生和发展[11]。本研究将采用分子对接和分子动力学模拟技术以及细胞实验研究BRAF蛋白在10-G抗黑色素瘤效果中的作用。
材 料 和 方 法
1 主要材料
10-姜酚中药对照品购自成都曼思特生物科技有限公司(MUST-17033004;批号:23513-15-7;纯度>99.04%)。DMEM培养基和胰蛋白酶购自Gibco;胎牛血清购自BI;MTS细胞活力检测试剂盒购自Promega;BCA蛋白定量试剂盒和ECL显影剂购自美国Thermo;抗p-BRAF (Ser445)、BRAF、p-MEK1/2(Ser217/221)、p-ERK1/2(Thr202/Tyr204)、p-P38(Thr180/Tyr182)和GAPDH抗体购自CST。
2 细胞培养
人皮肤黑色素瘤细胞株A375购自中国科学院细胞库,用含10%胎牛血清、青霉素(1×105U/L)和链霉素(100 mg/L)的DMEM培养基培养,在37 ℃、5% CO2及饱和湿度条件下的细胞培养箱中培养。倒置显微镜下观察细胞生长状态,细胞生长呈80%~90%融合状态时,以0.25%胰蛋白酶消化传代,隔日换液,每2~3 d传代 1 次。
3 方法
3.1MTS法检测细胞活力 调整细胞密度至5×107/L,接种于96 孔板中,每孔100 μL。在培养箱中培养24 h后,加入含有不同浓度(10、20和40 μmol/L)10-姜酚的培养基,每孔100 μL,并设空白对照组,每组设6个复孔,继续培养24 h。弃旧培养基,每孔加入100 μL 含MTS的工作液(0.1% FBS ∶MTS=100 μL ∶20 μL),37 ℃恒温箱内避光孵育120 min,在酶联免疫检测仪490 nm 波长处测量吸光度,上述实验重复3次。
3.2细胞计数 调整细胞密度至2×108/L,接种于6 孔板中,每孔1 mL。在培养箱中培养24 h后,加入含有不同浓度(10、20和40 μmol/L)10-姜酚的培养基,每孔1 mL,并设空白对照组,每组设3个复孔,继续培养24 h。弃旧培养基,PBS洗3次,加入0.75 mL 0.25%的胰酶消化,30 s 后加入0.75 mL完全培养基终止,收集细胞到EP管中,取100 μL细胞悬液加到900 μL完全培养基中,Millipore Scepter细胞计数仪检测细胞浓度,细胞数量=细胞浓度(cells/L)×0.0015 L×10(稀释比例)。
3.3分子对接 使用AutoDock软件分析10-G与BRAF之间的关系。从PDB数据库获得野生型BRAF(PDB ID:1UWH)和V600E突变型BRAF(PDB ID:1UWJ)的空间结构。从ZINC数据库取得10-G(ZINC ID:ZINC43009609)的3D分子结构。采用Autodock Vina软件进行分子对接模拟,在分子对接前去除原始空间结构中的水和配体。采用软件YASARA进行能量最小化。用BRAF与已知的BRAF抑制剂BAY 43-9006(索拉非尼,sorafenib)的结合位点预测10-G与BRAF之间的关系。将10-G与BRAF进行分子对接,采用结合能评估结合能力的大小。采用Pymol 2.7软件进行3D作图。
3.5Western Blot检测蛋白水平 60 mm 培养皿培养细胞,PBS 漂洗后收集细胞,RIPA 裂解液裂解,提取总蛋白,用BCA 蛋白定量试剂盒定量,加入上样缓冲液,10% SDS-PAGE 分离,转膜,脱脂奶粉封闭1 h,加入 I 抗4 ℃孵育过夜,1×TBST 液洗涤3 次,加入 II 抗,37 ℃孵育1 h,1×TBST 液洗涤3 次。ECL 化学发光试剂盒显色,胶片曝光。
4 统计学处理
采用SPSS 19.0统计软件进行分析。实验数据采用均数±标准差(mean±SD)表示。重复测量数据比较采用重复测量方差分析;多组比较采用单因素方差分析。若具有方差齐性,则各组间多重比较采用LSD法;若方差不齐,则采用Welch稳健估计,再采用Dunnett’s T3方法两两比较。以P<0.05为差异有统计学意义。
结 果
1 10-G抑制A375细胞活力
细胞活力检测结果显示,10-G剂量依赖性地降低黑色素瘤细胞的活力,与对照组比较差异有统计学显著性(P<0.01),见图1A。细胞计数结果示10-G能剂量依赖性地降低A375细胞数量,与对照组比较差异有统计学意义(P<0.01),见图1B。
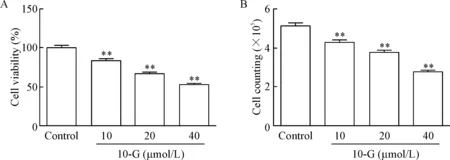
Figure 1.Treatment with 10-G dose-dependently inhibited the viability of A375 cells. A: cell viability (n=6); B: cell counting (n=3). Mean±SD. ** P<0.01 vs control group.
2 10-G与BRAF的分子对接
3 10-G与BRAFV600E复合物的分子动力学模拟
我们进一步采用分子动力学模拟技术探索BRAFV600E与10-G结合的稳定性。BRAFV600E/10-G复合物的表面模拟模型见图3A,左侧为分子动力学模拟初始模型,右侧为第100 ns的结合模型。经过100 ns的分子动力学模拟,10-G仍位于BRAFV600E的抑制剂结合位点。图3B展示了重原子均方根偏差root-mean-square deviation,RMSD)的变化轨迹。BRAFV600E/10-G复合物及非结合态BRAFV600E的RMSD均围绕3 Å波动。在最后20 ns,BRAFV600E/10-G复合物的RMSD稳定地低于非结合态BRAFV600E。

Figure 3.Molecular dynamics simulation of BRAFV600E/10G complex. A: binding conformation of BRAFV600E/10G at 0 ns and 100 ns; B: the root-mean-square diviation (RMSD) of heavy atoms.
4 10-姜酚抑制黑色素瘤细胞BRAF、MEK和P38磷酸化水平
我们将10-G(10、20和40 μmol/L)处理黑色素瘤A375细胞3 h后,Western blot检测p-BRAF、总BRAF、p-MEK1/2、p-ERK1/2、p-P38和GAPDH的蛋白水平。中、高剂量的10-G显著降低黑色素瘤细胞p-BRAF的蛋白水平,与对照组比较差异有统计学意义(P<0.01),低剂量的10-G对BRAF的磷酸化没有影响。低、中、高剂量组的10-G均能显著抑制MEK1/2和p-P38的磷酸化,与对照组比较差异有统计学意义(P<0.01),但对ERK1/2的蛋白水平没有影响,见图4。
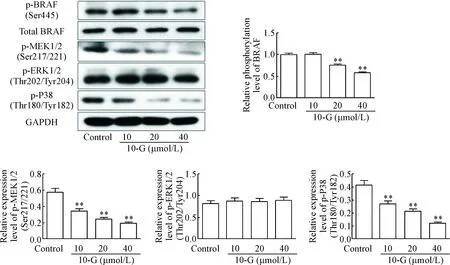
Figure 4.10-G inhibited the activation of BRAF, MEK and P38. Mean±SD. n=3.** P<0.01 vs control group.
讨 论
姜酚是造成姜的辛辣味道的主要物质,是由含有β-羟基酮结构的烷基链同系物组成的酚类化合物[12]。现代药物学关于姜酚的药理作用研究发现,姜酚具有降血糖、血脂[13],抗氧化、抗炎[14]以及抗肿瘤功能[15]。姜酚类化合物的抗肿瘤功能研究主要集中于6-G和8-G,关于10-G抗肿瘤作用的研究较少。目前仅在乳腺癌[15]和宫颈癌[16]上进行过10-G的抗肿瘤研究。黑色素瘤是最常见的皮肤肿瘤,恶性程度较高,是引起死亡人数最多的皮肤肿瘤,严重威胁人类健康。本项目对10-G的抗黑色素瘤作用进行了初步的探索。结果显示10-G能剂量依赖性地降低黑色素瘤A375细胞的细胞活力,减少细胞数量。这些结果提示,10-G具有抗黑色素瘤作用。
BRAF位于丝裂原活化蛋白激酶(mitogen activated protein kinase,MAPK)通路上游,具有参与细胞的增殖、迁移和分化的作用,其上游受到RAS的调控[17]。V600E突变是BRAF在黑色素瘤中主要的突变类型,BRAFV600E的活性是野生型BRAF的10倍[18]。更重要的是V600E突变的BRAF能不依赖RAS信号持续激活,进而激活下游信号通路,促进肿瘤血管生成,促进肿瘤细胞的转移,下调肿瘤细胞的凋亡反应以及机体的免疫反应[11]。因此抑制BRAF的激活特别是BRAFV600E的激活,对于治疗黑色素瘤具有重要意义。针对BRAF的特异性抑制剂具有良好的抗肿瘤药物开发前景[17]。
近年来,分子对接及分子动力学模拟技术广泛应用于药物研发及筛选中,可以显著提高药物开发的精准度和效率[19]。本研究通过分子对接模拟技术预测10-G与BRAFWT或BRAFV600E进行对接来预测复合物模型,分析表明10-G 与BRAF及BRAFV600E之间都具有较强的结合活性。我们还发现10-G与BRAFV600E的结合能低于10-G与BRAFWT的结合能(结合能越低提示结合能力越强),这提示10-G与突变型BRAF的结合能力高于野生型BRAF。我们还进一步采用了分子动力学模拟技术对10-G与BRAFV600E结合的稳定性进行评价,结果显示10-G与BRAFV600E的结合是稳定的。本研究采用BRAF抑制剂索拉非尼与BRAF蛋白的结合位点预测10-G与BRAF的结合位点。索拉非尼是2型BRAF抑制剂,能与BRAF激酶区的ATP结合域结合,通过与Cys531形成氢键促使索拉非尼占领BRAF的ATP结合域,还与Glu500和Asp593形成氢键稳定BRAF非活性构象[20]。本研究发现10-G能与Cys531和Glu500形成氢键,这与索拉非尼一致,MD模型也证实了10-G能稳定BRAF构象,这提示10-G的药理学机制可能与索拉非尼接近,是一种潜在的BRAF抑制剂。
为了进一步验证10-G与BRAF之间的关系,我们观察了10-G对黑色素瘤A375细胞BRAF激活的影响。结果显示,10-G能显著抑制BRAF的Ser445磷酸化水平。BRAF的Ser445磷酸化是BRAF激酶活化的标志,RAS激酶能磷酸化Ser445抑制BRAF激酶的自我抑制(auto-inhibition)使得BRAF持续激活[21]。BRAF抑制剂能稳定BRAF的非活性构象,使该构象的BRAF活化环(activation loop,A-loop)处于关闭状态,影响了上游的RAS对Ser-445的磷酸化,进而抑制了BRAF的激活[21-22]。我们推测10-G可能通过稳定BRAF非活性结构,影响RAS对BRAF Ser445位点的磷酸化。上述结果证实10-G具有抑制BRAF激活的作用,该机制可能在10-G的抗黑色素瘤过程中的发挥重要作用。此外,BRAF V600E突变已被证实在胶质瘤[23]、直结肠癌[24]和甲状腺癌[25]中起了重要作用。这提示10-G可能具有防治这些肿瘤的潜力。
本研究中我们还进一步探索了10-G对BRAF下游的MEK和ERK激酶激活的影响。结果显示BRAF能剂量依赖性地抑制MEK1/2的磷酸化,但是不能影响ERK1/2的磷酸化水平。既往研究发现10-G在结肠癌上通过激活ERK1/2诱导了结肠癌的凋亡[26]。我们推测10-G可能通过未知的途径上调了ERK1/2的磷酸化水平,抵消了抑制MEK1/2对ERK1/2活性的影响。我们进一步检索了STRING蛋白质相互作用数据库,发现MEK具有调控P38活性的作用。因此我们进一步检测了10-G对P38激酶磷酸化的影响,并发现10-G能显著抑制黑色素瘤P38的磷酸化。基于以上结果我们推测10-G可能通过抑制MEK而抑制P38的激活,从而发挥抗黑色素瘤作用。
综上所述,在本研究中,我们明确了10-G能抑制黑色素瘤的细胞活力,同时发现10-G可能是一个潜在的突变型BRAF抑制剂,其抗黑色素瘤作用与抑制BRAF密切相关。本次研究结果提示抑制BRAF可能是10-G及姜酚抗黑色素瘤的新靶点。
猜你喜欢
昆明医科大学学报(2022年8期)2022-07-31
波谱学杂志(2022年1期)2022-03-15
天津医科大学学报(2021年4期)2021-08-21
现代临床医学(2021年2期)2021-03-29
实用肿瘤学杂志(2020年6期)2020-12-09
天津医科大学学报(2019年6期)2019-08-13
祝您健康·文摘版(2019年1期)2019-05-14
天然产物研究与开发(2018年7期)2018-08-21
安徽医科大学学报(2015年9期)2015-12-16
安徽医科大学学报(2015年9期)2015-12-16
