
当归的亚高原区实验室培养与生长观察
2018-12-20 09:36李三相狄胜强赵菲佚王顺才
西北农业学报 2018年11期
贾 贞, 李三相, 狄胜强, 赵菲佚, 王顺才
(1.天水师范学院 生物工程与技术学院,陇东南植物功能基因组学与应用开发研究所, 甘肃天水 741000;2.山东农业大学 园艺学院,山东泰安 271018)
当归[Angelicasinensis(Oliv.) Diels]为伞形科多年生草本植物,主产于甘肃东南部的岷县等海拔为1 500~3 000 m的高寒阴湿地区[1],云南等地有少量栽培,其中,以岷县当归产量最多,质量最好,为当归传统道地产区[2]。成药后当归干燥的根为常用中药材,具有补血活血等功效,始载于《神农本草经》,除了药用价值外,当归还用于煲汤等食用领域,在中国当归栽培历史长达1 700多年[3]。当归生长周期长,从育苗、成药到种子生产,分3个阶段共3 a时间完成,第1年育苗,6月中旬播种,10月中旬起苗储藏;第2年成药,清明前后移栽,秋末采挖;第3年种子生产,选择长势好的成药当归根不收挖,留作第3年繁种结籽,8月中下旬采收。成药期部分当归抽薹开花而得不到具有药用价值的根,此即当归的早薹,严重时高达50%~90%[4-5],极大影响当归的产量。当归适应于高原低温区生长,海拔对当归早薹和长势均具有显著影响,株高、叶片数和抽薹率与海拔和经度均呈极显著负相关,株高、叶片数和抽薹率均与海拔间存在显著直线回归关系,可见,海拔是影响当归长势和早薹的主要生态因子,高海拔区种植的当归抽薹率显著降低,同时,偏低海拔区种植易于导致极低的产量[6]。朱国庆[7]对甘肃当归生态气候及适生种植区的区划分析显示,岷县、漳县及渭源南部海拔2 200~2 400 m间的气候冷凉阴湿半山区为最适宜种植区;临洮、渭源南部浅山区,海拔2 100~2 200 m间温凉湿润浅的高寒阴湿山区为适宜种植区;海拔2 000~2 100 m的岷县沿洮河河坝区及陇西南部温和湿润河谷山坝区、漳县东部川区;漳县、岷县、渭源南部海拔2 500~2 600 m的寒凉极湿润高山区,为次适宜种植区;漳县东部及岷县沿洮河海拔1 900~2 000 m 的温暖湿润川区,漳县、岷县及渭源南部海拔2 600~2 800 m的寒冷极湿润高山区,为可种植区;定西地区南部海拔<1 900 m或>2 800 m的地区,甘肃中部兰州、白银、临夏和甘南等地(州、市)及定西地区北部地区为不宜种植区,当归在该区大部分地方仍能成活,但气候条件及土壤质地不适宜当归生长,不宜种植,可见,当归的适宜种植区极为严苛。
获取试验材料的难易对该物种的研究具有重要的意义,模式植物拟南芥、水稻等是常温植物,其培养不受环境的限制,拟南芥的实验室培养方便,对拟南芥分子遗传学研究功不可没。因此,方便快捷地培养试验材料对一个物种的研究至关重要[8],但适应高原生境的药用植物当归则没有这么幸运,自明末栽培以来,当归始终而且只能在高海拔低温区栽培生长,当归育种和栽培的目标之一,是向偏低海拔区栽培引种,以扩大种植面积。其次,当归的适应特性极大限制了当归栽培范围及亚高原低海拔区实验室研究材料的获取,导致当归的实验室研究材料均取自当归产区岷县、渭源和临洮等地[9],不仅困难大,且存在取材不准、易受季节等的限制,给当归的实验室研究造成了极大不便,严重制约了当归的研究。本试验探索一种在亚高原区培养当归的方法,通过亚高原区室内培养,解决当归研究取材易受季节和气候性等的影响和限制的问题。试验使用当归种子、2 a生当归苗及成药后的当归根,在气温较高的亚高原城市天水市(平均海拔1 100 m,东经104°35′~106°44′,北纬34°05′~35°10′)实验室培养,探索通过实验室培养为当归研究供材的可行性,观察实验室培养中当归的生长及对亚高原高温的反应。
1 材料与方法
1.1 材 料
试验在2015-2017年间进行。试验所用当归种子获自甘肃省定西市岷县,带至实验室4 ℃冷藏,当归苗和成药当归宿根由岷县药农提供。当归盆栽使用规格为A300×200的花盆(外口径27 cm,内口径23 cm,高27 cm),花盆购自天水市花鸟市场,带地垫托盘。使用铭睿牌(E27型)4U-105W白光/黄色节能灯、E27平装式耐高温陶瓷灯座、户外悬挂防水灯罩遮光、正泰时控开关(KG316T)定时控制灯的开关和导线,1 m高铁架用于改装光照补光设施。在黑色塑料培养盘(宽30 cm,长50 cm,高15 cm)育苗。
1.2 方 法
1.2.1 当归苗和宿根的冷藏 当归苗可在前一年11月或当年3月带至实验室,依邱黛玉等[10]的方法进行苗的大小分类:用游标卡尺测量苗头下1 cm处的根直径,按苗直径大小分出直径小于0.45 mm的小苗和直径大于0.80 mm的大苗,保留没有机械损伤和病害者使用,暂时不用的苗捆扎用标签标示,用没有污染的湿土,储埋于合适大小的塑料盒内,置冰箱-5 ℃冷储,可用于当年或次年盆栽。
1.2.2 土壤的处理与盆装 盆栽所用土壤为多次栽培过大豆(Soybean)富含氮质的优质土壤,经砂网滤筛,搬至实验室,盛于大小合适的铁桶,置高温灭菌锅灭菌10 min,取出放凉,备用。处理好的土壤用于装盆,盆内不宜太满,与花盆边缘约2~3 cm即可,盆底垫托盘上浇水。
1.2.3 栽苗 苗的栽植方法与产区药农栽植方法一致:用小铲在盆内土壤掏出适宜大小的坑,将苗竖直放入坑内填土,苗头不宜高出土面。每盆栽4~6窝,0.45 mm以下的小苗2株/穴,苗根直径0.85 mm以上的大苗1株/穴,栽好后用保鲜膜覆盖盆口,以保温保湿,出苗后在膜上捅出部分小孔换气、放苗。
1.2.4 培养当归生长的光温 利用户外悬挂防水灯罩,配合E27型4U-105W白光和黄色节能灯(2个节能灯/灯罩)补光,灯高1.5 m(2灯/m2,1黄灯/2 m2),光照时长为16 h/d(4:00-20:00)。在室内自然温度下培养,反季节培养,借助室内暖气补温。
1.2.5 当归的RNA提取 分别取育苗期苗的第3个莲座叶、冬季培养成药期、成籽期当归的薹顶节完全展开复叶,用Invitrogen公司的TRIZOL试剂(cat#:15596026)提取RNA。使用TAKARA公司的反转录相关试剂,体系如下,10 μL变性液:Oligo dT Primer (50 μmol/L) 1 μL、dNTP Mixture (10 mmol/L each) 1 μL、模板RNA Total RNA 3~4 μg、RNase free H2O 定容至10 μL,94 ℃ 10 min变性、冷却。上述模板变性液、5×PrimeScript Ⅱ Buffer (4 μL)、PrimeScript Ⅱ RTase (200 U/μL) 1 μL、RNase Inhibitor (40 U/μL) 0.5 μL、无RNase H2O调节总体系至20 μL。反转录产物的验证和内标半定量使用当归肌动蛋白基因actin,当归开花关键基因AsFTL(AngelicasinensisFlowering locus T-like)用以表达验证,其引物如下(表1),55 ℃退火,内标基因Asactin的PCR扩增调整至30个循环,AsFTL的PCR扩增为25个循环。
栽植后每隔7 d进行当归复叶叶片数的计数。数据整理使用Excel软件,数据分析使用DPS 7.05数据处理系统[11],用Origin 6.0软件绘图。
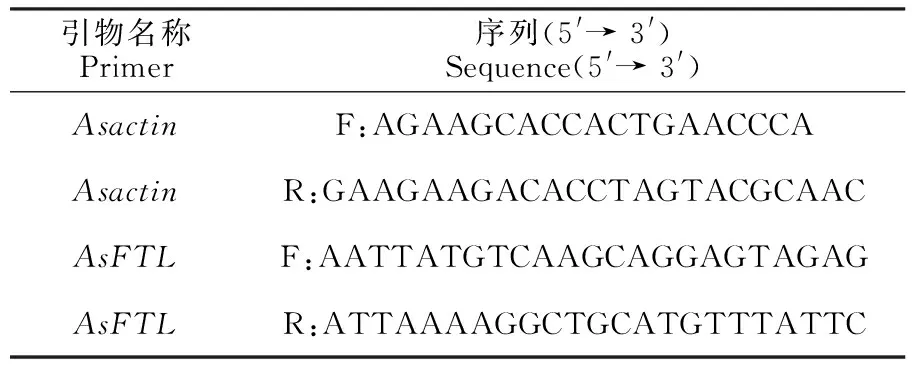
表1 开花基因表达检测用引物及其序列Table 1 Agronomic traits of dormant medicine-forming Angelica roots induced by summer hot
2 结果与分析
2.1 当归的实验室育苗
当归的室内育苗采用预先萌发,然后点播的方法。种子预先在培养皿中润湿(2017-04-01),置室温萌发12 d,萌发露白(图1-A)。挑选萌发的种子点播至盛满土的培养盘内,株间距1~2 cm,保鲜膜覆盖保湿,挑播的当归苗成活率接近100%,生长良好,自萌发后60 d长出2片真叶复叶,后缓慢增多,叶面积不断增大,至 102 d时达3片复叶(图1-B)。小喷壶随时喷水以使土壤和苗始终处于润湿状态,3~6月苗生长良好。
2.2 成药期当归的实验室培养与生长观察
成药期当归的培养每年分冬春季2个季节进行,春季培养既可用-5 ℃冷储当归苗,也可采用当年从产区带至实验室的苗盆栽, 9月反季节盆栽使用冰箱冷储苗。
2.2.1 成药期当归的春季培养 成药期当归的春季培养与生长:从4月开始,当归苗定期分2次盆栽,第1批4月9日盆栽,9 d后长出1~2片复叶,此后,叶片数逐渐增多,叶面积逐渐增大, 14 d 时小苗当归的叶片长度和叶面积明显小于大苗当归(图1-C),生长至48 d时,大苗当归出现抽薹(图1-D),此后,薹日渐增高,节数增多,至56 d时,薹上长出饱满的花序苞,当归的伞型花序即将进入初花期(图1-D 蓝框);未抽薹当归叶片数不断增多,叶冠增大(图1-E)。随夏季气温的升高,栽苗后95 d时各类当归苗休眠(图1-F)。从春季培养成药当归的生长动态来看,盆栽75 d内,始终保持良好的生长势头,叶片数日渐增多,至热休眠前,未抽薹当归最多可长至8个复叶,晚2月的第2批盆栽苗,因高温的胁迫,长至3、4个叶片后即休眠(图2-A)。
成药期当归的热休眠反应:亚高原区当归的实验室培养最难熬的是夏季高温,6月前能正常生长,6月底至7月初,待植株生长至7、8个莲座叶时,随气温的升高,叶片逐渐变黄、枯萎,但根仍然存活,即进入“夏休眠”状态。分析显示,休眠当归根的主根长与早薹当归主根长度间差异不显著,早薹当归的主根直径、侧根数、鲜根质量均显著高于未早薹的当归(P<0.05)(图1-F;表2),可见,成药当归的生长早期,早薹植株活力旺盛,生长能力强,形成较高的生物量,根和地上部分的一些性状显著高于未抽薹当归的相应性状,从二者地上部分的生长上也可以观察到这一现象。早薹当归抽薹进入花期后,其根丧失了进一步生长的能力,但未抽薹当归的壮根生长主要在秋季短日低温下进行,并形成强壮的药用根。
2.1.2 成药期当归的反季节培养 当归苗在岷县按传统方式贮存越冬,3月带至实验室-5 ℃冷储,于9月11日盆栽。成药当归的越冬期反季节培养也获得良好生长,栽后65 d各类当归均处于营养生长或营养体生长期(图1-G),栽苗后72 d大苗当归薹上第1节显现,即进入抽薹开花的生殖生长期(图1-G 蓝框)。成药当归的反季节培养中,110 d时未抽薹当归长出7个莲座叶。晚栽2月的同期第2批当归也能正常生长,至试验结束时35 d内长出3个复叶。成药期早薹当归长出4~5个莲座叶抽薹,该过程约需2个月,待完成抽薹约需110 d,此时,当归已长至第7个莲座叶,晚栽2月的当归,也能顺利成长,在试验结束时,1月内长出3个莲座叶(图2-B)。可见,无论早薹或未抽薹的营养生长时间,均显著长于成药当归春季培养生长发育时间,这可能与冬季气温较低有关。
2.3 成籽期当归的实验室培养与结籽
收挖2 a生当归的成药根, 在10月底当归成药根的收挖期,挑选根体积小、未受虫害或机械损伤的根,带至实验室,置冰箱-5 ℃冷藏,次年9月取出反季节盆栽。成籽期当归的培养在春季和越冬期各进行1次,春季培养在3月18日栽,反季节培养中,2 a生当归成药根经约1 a的冻储,次年9月初取出冷冻室置4 ℃冷藏萌芽,于2016-09-23盆栽,盆栽时2 a生当归的根仍然保持良好的存活状态,仅个别根毛处现少量白色发霉(图1-H)。成籽期当归的生长速度较快,栽后20 d已长出4片复叶,第4片复叶是后来薹上的第1复叶,栽后37 d出现薹上的第1节(图1-I,图3),栽后124 d长成完整结籽当归伞型花序,籽粒生长发育良好。成籽期当归栽后40 d完成主薹上全部叶片的生长,其中3个莲座叶、薹上3个主茎节及上端主茎节均可长出侧枝,形成侧生伞型花序(图1-J,图3),成籽当归生长后期开花结籽较慢,成籽约需要130 d。春季培养和反季节培养生长动态相似(图3),春季培养中因后期的夏季高温,难以形成有效的种子。在冬季反季节培养中,足够的低温和时间足以完成种子的生长成熟。

A-B. 当归的育苗 Photos ofAngelicaseedling;A.当归种子的萌发(4月13日) Germination ofAngelicaseeds cultured on dish for 12 d (April 13th shoot); B.培育102 d的当归苗(7月12日),白框内为相应时期的放大Angelicaseedlings cultured for 102 d (July 12), it is enlarged in the white frame;C-G.期当归的成药培养(2016-04-09栽) Photo series of medicine-formingAngelica(planted on April 9, 2016);C.栽苗后14 d(4月23日),黄色三角指示当归苗类的大小 YoungAngelicaat 14 d AP (April 23),yellow triangle indicates the size ofAngelicaseedlings;D.栽后48 d的抽薹苗(5月27日) Bolt appeared on 48 d AP(May 27);白框内为抽薹当归后期形成的由花序鞘包裹的花序苞 The inflorescence at 65 d AP is shown in the frame of the earliest boltingAngelicabefore flower open;E. 栽苗后56 d的未抽薹营养生长当归(6月4日) The vegetativeAngelicaat 56 d AP (June 4th);F. 休眠当归的收挖根 The harvested roots of the dormantAngelica:1、2为未抽薹营养生长当归的根 1, 2 are the vegetative roots,3、4为早薹当归的根 3, 4 are the boltedAngelicaroots;G. 栽苗后72 d的冬季反季节培养未抽薹营养生长当归(2016-09-11栽,11月22日拍照) VegetativeAngelica(planted on September 11, 2016) cultured in winter season at 65 d AP (November 22);白框内为栽后65 d的早薹当归(11月6日拍照),黄色箭头所指为首节 The boltingAngelicaat 72 d AP (November 6) is shown in the frame;the yellow arrow points to the first nod in bolt;H-J.3 a生成籽期当归的成籽培养 Photographs of triennialAngelicain winter growth;H. 经冬储的2 a生成药当归根(2016-09-23) BiennialAngelicaroot overwintered ice box storage (September 23, 2016);a. 1 a生当归小苗 Small seedlings are for medicine-forming growth;b.1 a生当归大苗 Bigger seedlings are for medicine-forming growth;c. 2 a生成药当归根 Biennial root seedlings are for reproductive growth for seeds;I. 栽后20 d(2016-10-12) TriennialAngelicaat 20 d after plantation (October 12, 2016);白框内为栽后37 d出现的首节(2016-10-28) TriennialAngelicaat 37 d AP, and the first nod of bolt appears (October 28, 2016);J.栽后124 d的成籽期当归(2017-01-24拍照),白框为花序的放大 TriennialAngelicaat 124 d AP (January 24, 2017), the amplified inflorescence is shown by the white frame;AP.after plantation

图1 育苗、成药和成籽期培养当归的生长发育Fig.1 Cultured Angelica at seedling, medicine-forming and seed-producing stages
注:苗于2016-03-18盆栽,7月3日热休眠收挖。数值为“平均数±标准误”。
Note:Seedlings were potted on March 18, 2016, and the roots were collected on July 3.Value of seedlings was “mean±SE”.
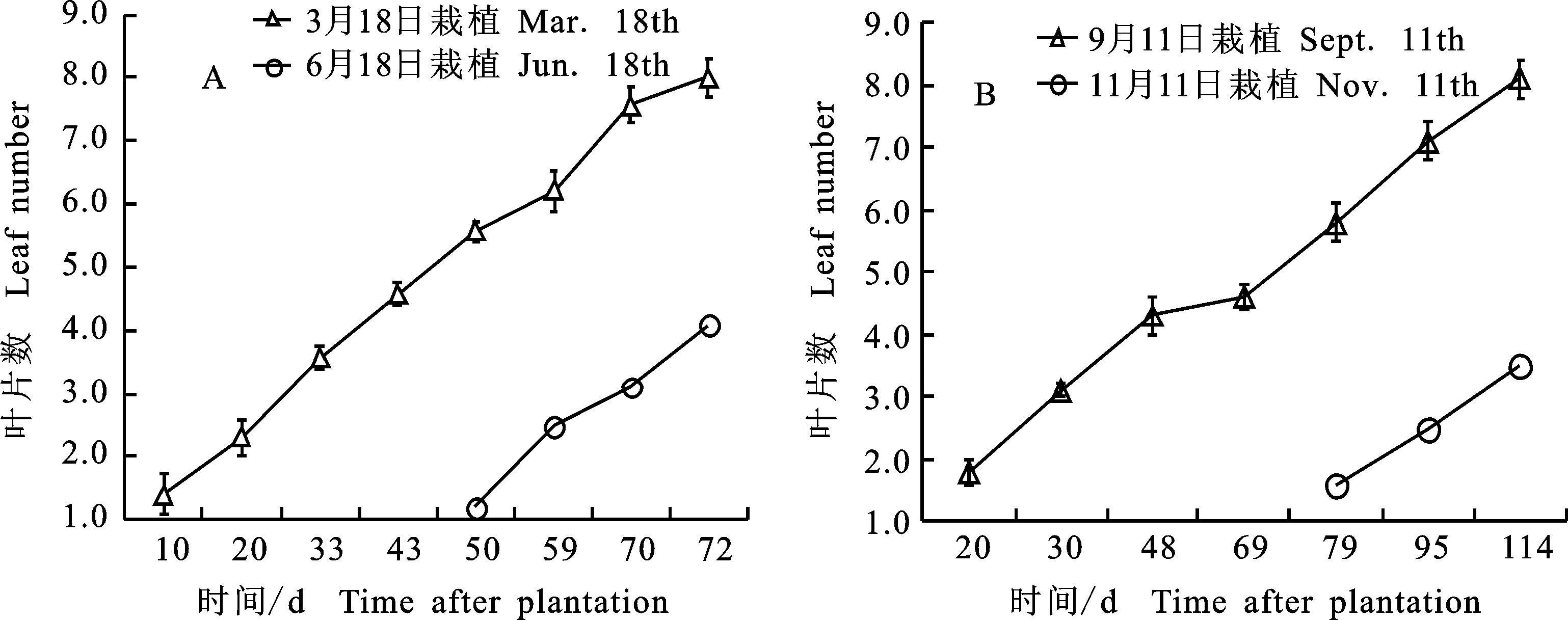
A. 春季培养,分别栽于4月9日、6月9日 In spring culture, the seedlings was transplanted on April 9 and June 9,respectively;B. 反季节培养,分别栽于9月11日、11月11日 In overwintering culture, the seedlings was transplanted on September 11 and November 11 respectively;各值为3盆共15株苗的“平均数±标准误” The mean was more than 15 seedlings in 3 pots,“mean±SE”
图2实验室培养成药期当归的生长动态
Fig.2Growthdynamicsofculturedmedicine-formingAngelica
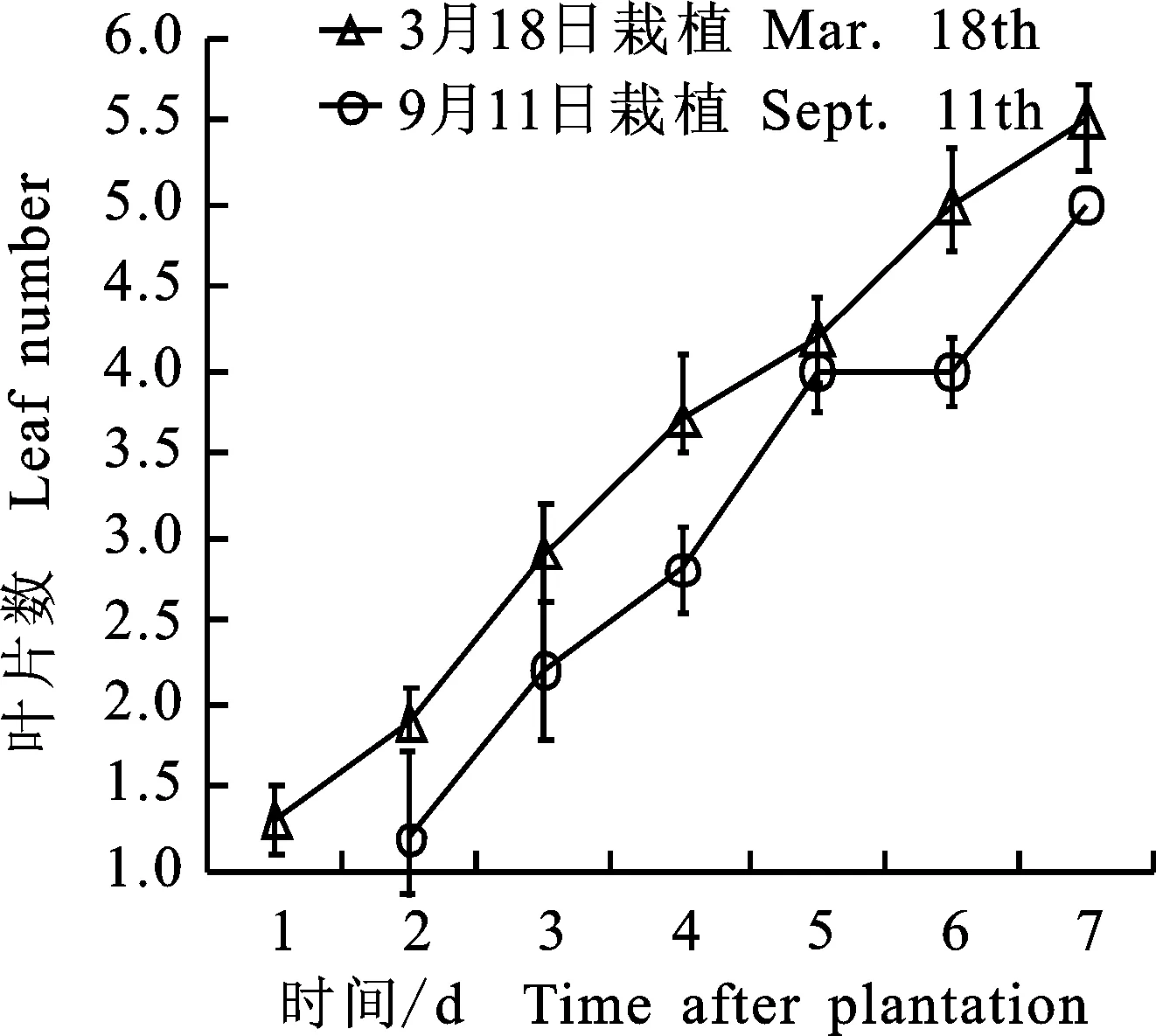
各值为3盆共6个植株的“平均数±标准误” Mean was more than 6 seedlings in 3 pots “mean±SE”
图32a生成籽期当归的生长动态
Fig.3GrowthdynamicsofAngelicaforseedsinthethirdyear
2.4 当归AsFTL基因表达的RT-PCR半定量检测验证
取育苗期完全展开的第3复叶、冬季培养成药期和成籽期当归薹顶节上完全展开的复叶,提取RNA,反转录后做当归重要开花基因AsFTL(GenBank: MF795555)表达的RT-PCR半定量。结果显示,在各时期当归叶片内参基因Asactin亮度接近的情况下,育苗期的当归叶片内AsFTL极为微弱或检测不到,但在成药期早薹和成籽期当归的叶片中,AsFTL的表达量较高,二者表达量接近(图4),显示,AsFTL的表达与当归抽薹开花有关。
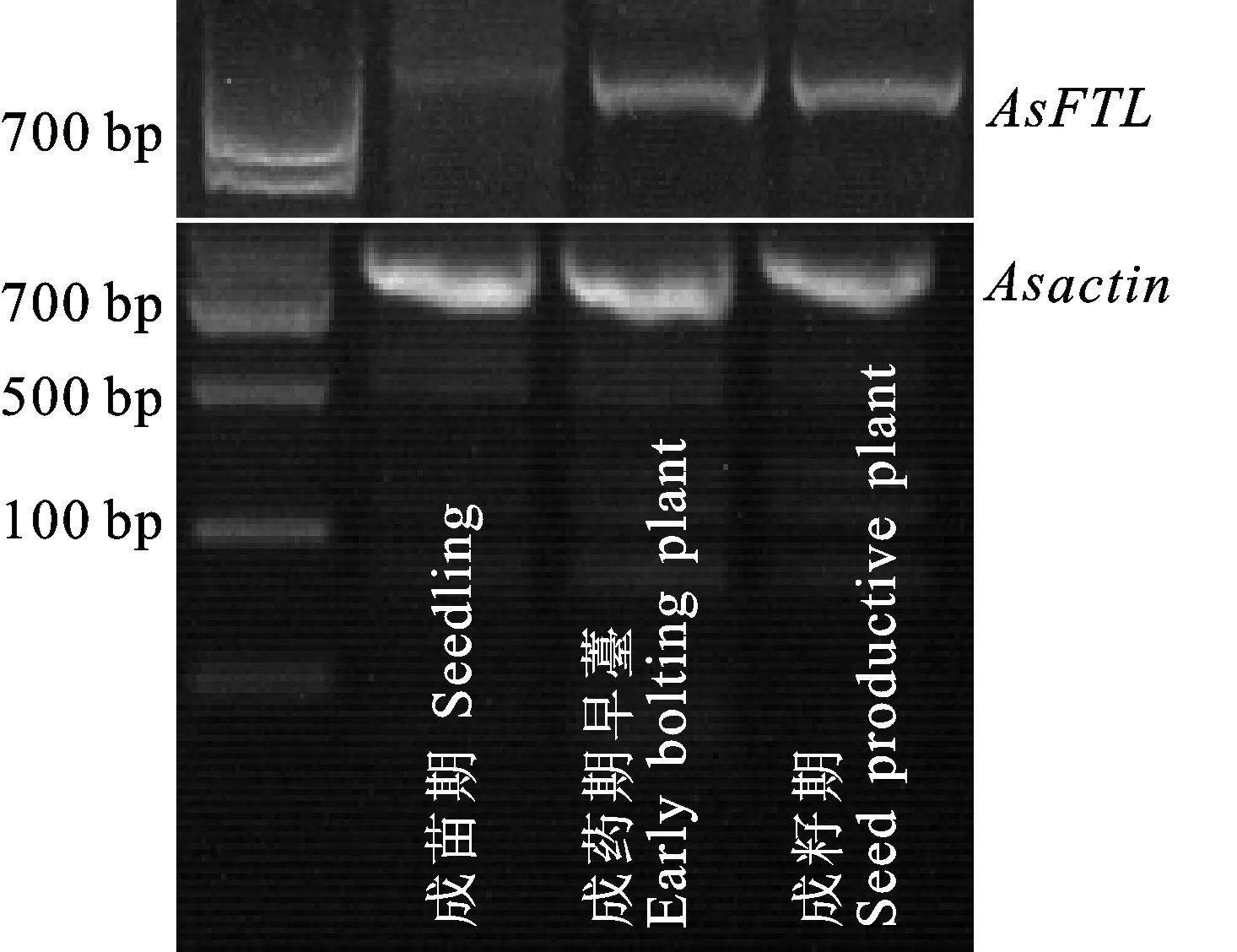
图4 当归叶片AsFTL表达的RT-PCR半定量检测Fig.4 RT-PCR semi-quantitative detection of AsFTL expression in leaves of Angelica
3 讨论与结论
在道地产区,当归种子于夏至播种于海拔约3 000 m以上的生荒地,本研究在亚高原区实验室于4月进行播种育苗,从种子萌发到苗的生长,在6月前气温低,苗的生长良好,但随夏季气温的升高,苗面临高温胁迫,这种情况下,若能辅佐以空调降温则可维持苗的生长。当归的反季节育苗已有较大进展[12],实验室同样可进行当归的反季节育苗。研究中当归的春、冬季成药培养均能正常生长,遵循大苗抽薹小苗营养生长的常规现象,但春季培养因气温高而生长快。阳坡、低海拔高温[5,10]下栽培易导致当归早薹,因此,对当归来说上述高温是一种逆境,在该逆境下,当归以早薹开花结籽的方式缩短生活史,以回避逆境[13],是植物对高温胁迫的逃逆反应[14]。本研究中未早薹当归则进入夏休眠状态,说明,当归的高温反应除早薹开花外,未能抽薹的小苗当归的高温反应则是休眠。温度是诱导植物开花的一个因子[15-16],通过温度调控途径实现[17]。在适度高温诱导下,具有早薹潜力的大苗当归进入抽薹开花和结籽等生殖活动以保存后代,而没有早薹潜能的小苗当归则仍维持营养生长,在更高的温度下,这些当归因不能持续生长而休眠。因此,笔者认为早薹是当归对适度高温的反应,而在更高温度下,则发生当归根的休眠,但后者仅限于未抽薹的营养生长当归。夏休眠的诱导及生长由活跃期向停滞期的转变涉及重大的发育变化,如休眠再生芽的形成、分生组织活动的停止,导致叶发生的停止及随后地上部分的衰老以及代谢向储藏器官的再转移[18],长日高温是一些植物夏休眠的诱因[19],ABA可能参与这一过程[20]。当归的早薹或高温下休眠反应限制了当归的有效栽培范围,随全球气温的变暖[21],当归的种植范围不仅难于扩大,缩小也在情理之中[22]。由此,当归可在亚高原区实验室培养,在面对夏季高温会做出逃逸反应,但当归的越冬反季节培养则不存在这一问题。本研究中当归的育苗和成药培养自4月开始,与道地产区当归的成药栽植同步,但依亚高原区的气温,当归的实验室培养完全可提前至2月3月进行,最晚可在5月进行,冬季反季节培养待气温转凉的9月初即可进行。3 a生成籽期当归根的收集尽可能选用最小的根,以减少储藏空间。FT(Flowering Locus T)及其同源基因是植物开花调控途径的重要整合因子和调控开花的关键基因之一[23],已被证实是拟南芥等多种植物可传导开花物质“成花素”的编码基因[24]。FT的同源基因的表达可缩短苹果和柑橘等多年生植物的成花年限[25-26]。当归AsFT基因(AngelicasinensisFlowering Locus T)及其同源基因AsFTL是当归早薹开花的关键基因。本研究中通过检测不同发育阶段当归叶片AsFTL的表达,以验证培养当归做为试验材料的可行性。半定量RT-PCR分析显示,育苗期当归中没有AsFTL的表达,而在成药期早薹当归和成籽期当归叶片中存在强烈表达,这与其他植物FTL同源基因及笔者关于当归早薹相关转录组分析结果一致,说明亚高原实验室培养的当归可用于其基因表达研究。
温度、光照和土壤是影响当归室内培养的主要因素。(1)温度:当归生长对低温有很强的依赖性,道地产区岷县及周边平均海拔为2 200~2 400 mm[27],对比岷县和天水市秦州区气温变化,天水市秦州区7月、8月白天气温可超过30 ℃,达35 ℃,夜间最低气温也在20 ℃左右,昼夜温差小,此时岷县白天最高气温接近或偶尔高于30 ℃,夜间气温可低至15 ℃以下,昼夜温差大,而大田实际温度更低,是适合当归生长的温周期。温周期通过调节植物内源细胞分裂素改变植株昼夜间硝酸还原酶的表达和活性的循环模式[28],对植物的种子萌发[29]等生长发育具有十分重要的意义。当归的实验室春季培养符合当归的温周期,白天气温高,当归生长快,而反季节培养因依赖于室内暖气,夜间温度较高,不符合当归的温周期,但也能正常生长,若能在夜间辅以空调等降温设施则更符合当归的生长特性。(2)土壤:当归是大田作物,生长周期长,成籽期当归具有硕大的根,盆栽培养盆不宜太小,蛭石等基质也不适于当归的培养,否则会造成后期营养缺乏。当归的盆栽培养只能使用土壤,产区当归的栽培以肥沃、土层深厚的黑土或褐土类、质地疏松及排水良好的肥沃富含腐殖质的砂质土壤为佳,不宜在易板结的粘土和贫瘠的砂质土上栽种[7,30]。本研究中当归培养所用土壤为经多次栽培大豆(Glycinemax(L.) Merr)的黑色土壤。当归的根极易受土壤微生物的污染和虫害而腐烂,因此,土壤用前需经高压灭菌锅灭菌10 min,以杀死土壤内的线虫等爬虫类害虫和杂草类,减免对根苗的污染和苗期虫害,保证苗的健康生长,也可除去草籽消除杂草,保证盆内清洁。产区当归的育苗在“生荒地”进行,同地块经历一两次育苗后,10 a后方可进行下次育苗。本研究中当归的育苗使用与盆栽相同的普通土壤,亦能获得正常的生长苗。(3)光照:本研究中当归的培养通过节能灯补光,光照度为2 000~3 000 lx,完全满足当归的生长,光照的时长依试验需要设置,本研究使用16 h的长日光照,旨在促进当归生长和早薹。当归的室内培养平时还应注意苗期管理,当归性喜凉爽和湿润的环境,在空气湿度较大的环境下生长良好,因此,依盆内土壤干湿程度适时浇水,使土壤始终处于湿润状态。为防止盆面上浇水形成板结,采用在盆垫托盘上浇水的方法,每次浇水加满托盘。当归培养期间可用小喷雾器在植株地上部喷雾,以缓解环境气温的升高和保湿。
猜你喜欢
农产品市场周刊(2022年3期)2022-03-02
烟台果树(2021年2期)2021-07-21
学生天地(2020年18期)2020-08-25
饮食保健(2020年10期)2020-05-29
建材发展导向(2019年5期)2019-09-09
中国社区医师(2019年13期)2019-08-31
建材发展导向(2019年10期)2019-08-24
中国民族医药杂志(2016年5期)2016-05-09
中国火炬(2014年10期)2014-07-25
地震研究(2014年4期)2014-02-27
