
植物夜间液流的发生、生理意义及影响因素研究进展
2018-12-19 10:44:34方伟伟傅伯杰
生态学报 2018年21期
方伟伟,吕 楠,*,傅伯杰
1 中国科学院生态环境研究中心,城市与区域生态国家重点实验室,北京 100085 2 中国科学院大学,北京 100049
植物对水分的利用包括白天和夜间的蒸腾作用,目前已有大量的研究结果阐释了植物在白天对水分的利用过程[1-2],但是植物在夜间对水分的利用过程研究较少。植物的夜间液流是指在夜间土壤中的水分进入植物的根系后,通过输导组织向上运送到达冠层,或者储存在植物的茎干部分,或者通过气孔蒸散到空气中。之前有研究认为植物在夜间气孔是处于关闭状态[3],不存在夜间液流现象。后来随着观测技术的进步与发展,研究人员通过各种观测手段和技术,对植物不同尺度、不同生境的物种的夜间液流进行了观测,结果表明几乎所有的植物均存在夜间液流现象[4]。夜间液流的普遍存在,使得学者开始对它存在的意义进行探讨。由于夜间液流只有水分的损失而无碳的固定,所以早期的研究认为夜间液流是纯粹耗水的过程,不利于植物的生长。随着研究的深入,研究者发现夜间液流不仅包括夜间蒸腾过程,还包括植物茎干的储水过程。茎干储水过程对植物第二天的光合作用、蒸腾作用以及抗干旱胁迫有重要生理意义;而夜间蒸腾过程对植物的营养物质和氧气运输等方面有重要作用。这些研究结果说明了夜间液流的发生并不是偶然现象。
在生态水文模型中,由于夜间液流占全天液流的比例相对较少,所以传统的生态水文模型很少考虑或者直接忽略了夜间液流的影响。但是,对于某些物种和某些地区中,夜间液流可能占全天液流很大的比例:如降雨量较少的地中海气候类型区,冬青栎(Quercusilex)可达40%[5];美国中部的油栎(Quercusoleoides)可由充沛降雨条件下的8%上升到干旱条件下的20%[6]。夜间部分液流用于蒸腾,则用模型计算生态系统的水分收支时就会造成误差,水量将会低于实际值[7-8],进而影响人们对水资源的管理和决策。为了减小误差,也需要对夜间液流进行准确的定量分析。有研究表明未来气候变化会使得夜间温度等环境条件发生变化,这将会影响到夜间液流过程以及夜间液流在蒸腾与茎干储水间的分配,使得夜间蒸腾对水量平衡影响的不确定性也更大。
环境因子通过影响夜间液流而影响着植物的生理过程,进而影响着植物对环境的适应,同时部分夜间液流也通过夜间蒸腾而影响着样地水量平衡。虽然研究者已经对夜间液流(气孔导度)进行了观测,但是关于夜间液流存在的生理意义和影响因素等方面还没有全面的了解,关于夜间液流的观测方法,以及各种方法的运用尺度和优缺点已有学者进行了总结和归纳[9-10],因此本文在总结国内外已有研究的基础上,从夜间液流现象的发生、组成、生理意义、影响因素以及对生态水文的影响等几个方面对夜间液流的研究现状进行阐述,并针对气候变化条件下夜间液流的响应,提出今后的研究方向。
1 夜间液流的发生及大小
通过对不同生境(如热带雨林[11-12]、温带常绿阔叶林[13]、半干旱区[14-15]、沙漠[16])的乔灌丛,不同生活型的物种(一年生和多年生草本[17-18]、灌木[19]、乔木[20]),不同的生态系统(天然生态系统[17, 20],人工生态系统[21])等的夜间液流进行观测,发现几乎所有的物种均存在夜间液流现象,但是存在量级的差异。为了方便与其他的研究做分析比较,文献中一般用夜间液流占全天液流或白天液流的比例来表示夜间液流大小。我们对已发表的文章进行了检索(检索数据库为:web of science;检索的关键词为:“nighttime sap flow/transpiration”,“nocturnal water use”,“nighttime stomatal”;检索年限:1980—2017),排除控制实验的文献,共收集收已发表的38篇文献中的数据(共有95个物种,包括51个属)(见附件)。结果发现植物夜间液流(Enight)占全天液流(Edaily)的比例一般为5%—25%(图1),如在半干旱区的黄土高原,灌木夜间液流一般占5%—20%[19],澳大利亚地区桉属植物(Eucalyptusforest)所占的比例为8%—10%[21],美国北卡罗来纳州的山核桃属(oak-hickory forest)可以达到22%[22]。但是在某些特定环境下,夜间液流所占的比例较高,如干旱区植物的夜间液流能够占到白天液流的60%[16]。在研究过程中发现有少数植物没有夜间液流现象或者夜间液流比例很少,如加利福尼亚州的鼠李(Ceanothuscuneatus)[20]、地中海气候区的热带稀疏草原的柳叶石楠(Heteromelesarbutifolia)[20]等。
植物的夜间液流速率受到气候条件、树龄和光合作用途径等多个因素的影响[8]。我们用收集到的95个物种的夜间液流占全天液流的比值,分析了不同植被类型下夜间液流占全天液流的比例,结果发现在热带雨林和热带稀疏林地区夜间液流比值要显著高于其他地区(P<0.05),但是大部分的研究是关于乔木的,而对灌木和草本的研究较少(图2)。对于同个物种来说,夜间液流所占比例也会受到叶片或者植株的年龄[23]、群落受干扰程度及恢复年限的影响。研究结果表明年龄较小的叶片具有较高的夜间气孔导度[21];墨西哥地区林龄较小的人工松树林的夜间液流比例(42%)要高于成熟林龄的(19%)[4];受火干扰之后桉树林的夜间液流要比自然林的高10%[24]。受干扰(自然或人为)之后的群落为研究不同林龄之间夜间液流的差异提供了很好的自然条件,所以在以后的研究中,可以采用人工干预的方法来研究夜间液流与林龄的关系。夜间液流也与植物的光合途径有关,在C3途径和CAM途径之间可以转换的植物中有着较高的夜间液流[25]。同时我们也分析了不同的生活型(乔木、灌木和草本)之间夜间液流的差异,结果显示不同生活型之间的夜间液流的比例并无显著性差异(P>0.05)。这些结果表明夜间液流与植物的功能性状、生理特征等因素有关,但是生活型之间夜间液流的差异可能受到样本量的影响,以后的研究还需更多物种和地区间的对比。
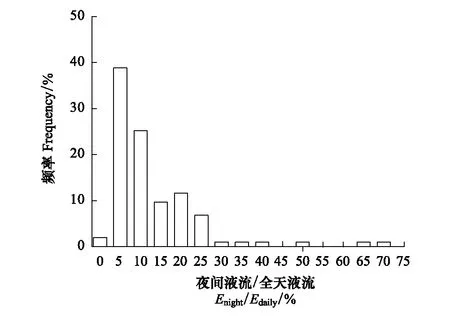
图1 夜间液流占白天液流发生的频率分布图 Fig.1 Frequency distribution of the percentage of nighttime sap flow to daily rates in 96 species
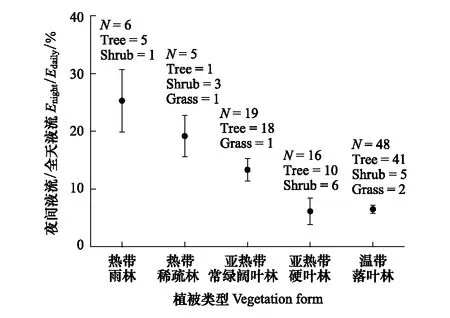
图2 不同植被类型植物夜间液流占全天液流的比例均值 Fig.2 Mean values of nighttime sap flow to daily rates for various vegetation types 在图中标明了不同植被类型下三种生活型的物种数,N:总物种数,Tree:乔木物种数,Shrub:灌木物种数,Grass:草本物种数
2 夜间液流的组成
夜间液流并不等同于夜间蒸腾。夜间液流一部分通过气孔发生蒸腾作用,另一部分用于茎干补水。两者发生的过程不同,受到的影响因素以及对植物的生理意义也存在差异。目前并没有确切的研究或技术方法将两个过程定量的区分,因为两个过程是同时发生或者是先后发生的,而且研究表明夜间蒸腾过程可以带动补水过程[26]。研究表明夜间的补水作用占到夜间液流的40%—70%,而夜间蒸腾占到30%—60%[27]。有学者对区分两个过程进行了初步的尝试。根据植物的蒸腾作用与水汽压亏缺的响应关系进行区分,因为引起夜间蒸腾的主要因素是水汽压亏缺,也就是当水汽压亏缺很小或为零时,如果液流依然存在,那么这部分液流是用于茎干补水的,如果液流与大汽压亏缺有很好的拟合关系,那么这部分液流主要是用于植物的蒸腾作用[7]。有研究通过液流速率对水汽亏缺的响应发现有些植物只有夜间蒸腾作用而无补水作用,如纸皮桦(BetulapapyriferaMarsh.)[7]。也有学者利用仪器进行监测,在样株的基部和顶部各加一个传感器,基部传感器的数值为总的夜间液流量,而通过顶部传感器的液流值则可认为是蒸腾量,两者的差值即为茎干补水量[13]。但是这两种方法均存在误差和限制条件:在第一种方法中由于影响夜间液流的因素是多样的,需要将所有的因素综合考虑再做分析;第二种方法对高大乔木操作不方便,一般适用于灌木或草本植物,而且这种方法仅仅考虑了茎干的储水量,没有考虑叶片的储水量。所以最好通过测定总的夜间液流量,再通过测定整个样株的蒸腾量来分别定量分析夜间液流的两个过程。
3 夜间液流的生理意义
3.1 茎干的补水作用
植物夜间液流的一部分用于茎干的补水[13,15]。有研究认为植物茎干的粗细与植物储水量有关[28-29],所以高大乔木的储水量要高于草灌植物的储水量。植物茎干的储水作用,提高了黎明前茎干和叶片的水势,从而使植物的气孔导度能在黎明前提前开启,提高第二天叶片的光合作用[30],避免因土壤水分或者光照条件等环境因素的限制而影响植物的光合作用[19, 20],植物的这种优势在干旱和半干旱地区尤为明显,使植物在一定程度上缓解了干旱条件对黎明前光合作用的影响,缓解了因干旱胁迫而影响植物的碳固定过程;同时也可以减少植物木质部栓塞化的形成[31],例如在热带稀树灌木林地的存在茎干储水作用[32]。由于植物存在着夜间的储水作用,第二天上午的蒸腾速率与前一天夜间液流速率呈现显著的相关性[33],使得第二天上午的蒸腾作用可以优先利用茎干部分的水分[33],促进光合作用,增加光合作用时间,从而促进植物生长[34]。在水分充足或湿润地区,由于没有水分的限制,植物的夜间补水过程是否存在,或者夜间补水占夜间液流比例及生理意义还没有具体的研究,所以植物茎干的补水作用是否在所有物种和生境中存在还需要更多的野外观测。
3.2 水力提升和再分配过程
土壤的水力提升(hydraulic lift)过程是指植物根系将土壤水分从水分含量较高、较深的土层中重新分配到较干旱、较浅层的土层中[35]。在干旱和半干旱地区,表层的土壤水分含量较低并不意味着植物受到干旱的胁迫,植物可以通过水力提升作用将深层的土壤水分提升到表层供植物吸收利用[36-37],有研究表明水力提升过程与植物的夜间液流相关[38]。水力提升作用促使了水分和营养物质在植物根系周围的运输[39],植物可以更好的吸收水分和营养物质,所以在较干旱的地区,水力提升作用较为明显。水力再分配过程是双向的,是干旱生境条件下植物内在的一种生存机制[4],但是这一过程与水力提升作用相反,有研究表明植物的夜间液流抑制了水分的再分配过程[38],而这种抑制作用甚至会在干旱季节降低植物的生产力。水力提升和再分配过程通过改善土壤水分的有效性和根系的活力影响植物水分关系[40]。由于这两个过程均受到根系分布及根系深度的影响,所以这两个过程在乔木和灌木中的作用较明显。但是目前总的来说这方面的研究还较少,所以夜间液流与水力提升和水力再分配的关系以及与生境条件、生活型的关系还需要进一步的研究。
3.3 植物的氧气供给作用
持续的夜间蒸腾作用同时也将氧气运输到植物木质部的薄壁细胞中,这维持了植物细胞对氧气的需求[7]。由于在不同的气候区的乔木或灌木的木质部细胞均存在对氧气的需求,所以夜间液流对氧气的运输作用主要存在生活型的差异,夜间液流的氧气供给作用对草本植物的作用可能不太明显,但是对于高大的乔木植物以及灌木较为重要。白天由于植物的蒸腾作用促使氧气在植物体内运输[41],但是由于夜间的蒸腾作用降低,蒸腾拉力较小,而且远距离的运输使得到达乔木或灌木上端时液流中氧气含量也相对较低,所以能够给植物呼吸作用提供的氧气也较少,所以需要提高夜间液流量来供应植物细胞的氧气需求[41]。但是夜间液流对植物的呼吸作用在之前的研究中涉及较少,主要原因是植物细胞的呼吸作用和液流中运输的氧气含量均难以测定,夜间液流对植物的呼吸作用还需进一步的研究和观测。
3.4 营养物质的运输
夜间液流有一部分用于植物的夜间蒸腾作用[13]。蒸腾作用带动了营养物质的运输,所以有学者探讨了植物的夜间液流与植物营养物质吸收之间的关系。植物生长必需的营养物质溶解在土壤溶液中,在根系水力提升的作用下,通过土壤溶液运移到植物根系附近,所以一些研究结果表明植物夜间液流[42-43]或者夜间气孔导度[16,31]与N元素有关;也有学者通过对16个物种夜间液流的研究表明植物的夜间行为对N元素的吸收和黎明前光合速率有不同的影响[44],由于植物吸收养分的各个过程均需土壤水分的参与,所以只有在土壤水分充足的地区,夜间液流才能促进营养的吸收[38]。但是也有研究表明植物的夜间行为与N的吸收无关[45-46]。这可能与植物的生境条件有关,在降雨充足而N元素缺乏的地区,夜间液流的增加有利于植物对N元素的吸收,在降雨充足而N元素非限制区,夜间液流可能主要用于植物的蒸腾作用,促进水分循环[27]。所以由于植物的生长习性以及对环境的适应性不同,导致夜间液流的作用不同[47]。
4 夜间液流发生的影响因素
4.1 气象因素对夜间液流的影响
影响夜间液流的主要气象因素有水汽压亏缺、温度、风速等[7,48],多数研究表明夜间液流的主要影响因素是水汽压亏缺,如对纸皮桦(BetulapapyriferaMarsh.)的研究结果表明,水汽压亏缺可解释夜间液流66%的变异[7]。在干旱环境中由于夜间较高的水汽压亏缺,夜间液流占全天液流的比例较高,甚至可达60%[16],所以生境条件影响夜间液流速率。同时温度和风速与夜间液流速率也呈现着正相关关系[49],在湿润的环境中,夜间液流与水汽压亏缺的拟合关系不如在较干旱地区的[27],所以在热带雨林地区,较高的夜间液流比例可能主要是受到较高的夜间温度的影响[8]。除此之外,Zeppel等的控制实验表明在干旱条件下空气中CO2浓度对夜间液流速率起到抑制作用,但是,在湿润条件下,升高CO2浓度有利于夜间液流的发生[49]。由于环境中多种因素的共同存在,这几个因素的综合作用对液流速率的影响目前尚不清楚,还需要进一步的研究。
此外,夜间液流和白天液流对水汽压亏缺的响应不同。白天植物的液流速率随着水汽压亏缺的增加而增加,但是当水汽压亏缺增加到一定程度时,植物为了减少体内水分的损失而减小气孔导度,植物的液流速率会保持不变甚至会降低[33],所以植物白天的液流速率对水汽压亏缺呈现非线性响应,这主要是发生在日中午的一段时间。但是对于夜间液流,由于存在茎干补水作用,使得夜间液流速率对水汽压亏缺的响应没有白天敏感[19,50],而且由于夜间水汽压亏缺较白天要小,达不到植物气孔关闭的阈值[7]。
4.2 土壤水分对夜间液流的影响
研究结果表明除水汽压亏缺之外,土壤水分也是影响夜间液流的主要因素[4,51],但是目前土壤水分对夜间液流的影响还没有一致的结果。在干旱胁迫下,植物夜间的气孔导度随土壤水分的降低而减小,夜间液流随之降低[6]。Zepple 等研究结果表明当水汽压亏缺保持恒定时,植物夜间液流随土壤水分的减少而降低。同时,在其他研究中发现夜间液流速率与土壤水分呈现正相关关系[44,52],这是因为在干旱条件下,植物叶片对水分的穿透能力减弱,降低了液流速率。也有研究表明夜间液流的大小与降雨之后的天数呈负相关关系[20]。在黄土高原地区研究发现夜间液流占全天的液流比例随土壤水分的减少而增加[19],这表明在受到土壤水分限制时,植物可增加夜间液流的比例来缓解干旱[6,19]。除此之外,不同层次的土壤水分对夜间液流速率也有不同的影响[13]。在全球气候变化背景下,夜间温度在增高的同时伴随着降雨量的减少[53],使得夜间水汽压亏缺的增大及土壤含水量的减少,可能导致夜间液流所占比例增大,所以研究影响夜间液流的影响因素有助于预测未来气候变化对液流速率的影响。
5 夜间液流的生态水文效应
夜间液流不仅发生在几乎所有的物种上,甚至占到全天液流的5%—60%,这对植物水分关系产生了一定的影响。在计算植物的个体、群落、生态系统甚至小流域的水量收支平衡时,需要考虑植物通过夜间蒸腾而损失的水分。很多水文模型认为在太阳辐射为零时,植物的耗水量也为零[54-55],或者假设夜间无水分的损耗,那么模型的结果可能与实际数值存在差异。虽然有模型尝试着将夜间的气孔导度添加到生态水文模型中[56],但是由于植物的夜间行为存在着不确定性,同时受到环境因子的影响及植物自身的调节等作用,所以用气孔导度来表示水通量是否合适还不确定,也增加了模型的不确定性。在全球气候变化条件下,北半球的夜间温度比白天温度增加速率快[53],同时夜间水汽压亏缺的增加将会影响整个生态系统的水量平衡。因此,如果不考虑植物夜间的耗水量,将会低估植物蒸腾对整个生态系统水量平衡的影响[10]。
在没有夜间液流的条件下,土壤和植物叶片的水势可以达到平衡状态,因此在很多的模型研究中,直接将黎明前的叶水势当作土壤的水势来处理,甚至在一些观测实验上也将黎明前水势认为是测定土壤水势的工具[57-58]。但是大量的研究结果表明植物夜间气孔并不完全关闭[17, 22],而且存在着夜间的蒸腾作用和茎干补水作用,这就打破了土壤-植物水势之间的平衡[59],使得叶片水势在黎明前高于土壤水势,这种现象在干旱和半干旱地区尤为明显。由于技术手段的发展,对夜间蒸腾作用和气孔导度的测定存在着一定的限制,所以在运用模型时有研究通过白天的气孔导度行为来推算夜间通过气孔导度而损失的水分。但是有研究结果表明在不同的环境和物种条件下,由于植物的水分利用策略[60],光合作用途径与反馈机制[61]、生理特征[62]等因素的差异,使得植物的气孔导度对蒸腾作用的控制作用在夜间和白天展现出不同模式,所以不能简单的用白天的气孔导度来推算夜间的水分蒸腾损失。为了提高模型的准确性,最好是对夜间的蒸腾作用进行自动观测,或者是通过观测夜间气孔导度与夜间蒸腾作用的关系,进而推算夜间通过蒸腾作用而损失的水分。
6 未来研究方向
目前,虽然对植物的夜间液流进行了很多研究,但是还有很多问题没有解释清楚或者需要进一步的研究,主要可以从以下几方面来进行探讨:
(1)夜间液流与植物适应性关系的研究
夜间植物的储水作用与蒸腾作用对植物的生理意义不同,影响着植物对环境的适应性,但是夜间液流的两个过程的区分一直是测定夜间液流中的重点和难点。如何准确区分两者的比例是需要解决的关键科学问题,同时也需要与各种技术手段相结合。气孔导度在夜间虽处于半开放状态,但气孔导度与夜间蒸腾作用的关系并不完全清楚,是否可以用夜间的气孔导度来表示夜间的蒸腾作用还需要进行细致的研究。在降雨量充沛的地区,植物是如何通过蒸腾作用运输氧气和营养物质;而在干旱和半干旱地区,水分是限制植物生长的主要因素,植物是如何通过夜间的水力提升作用来提高缓解干旱的,夜间储水量与土壤水分,干旱胁迫三者之间的定量关系如何,是否可以通过夜间液流与植物叶片(茎)在夜间的水势差来确定植物的干旱程度,这需要进行更进一步的研究。
(2)气候变化条件下,建立从叶片夜间蒸腾、个体和样地尺度的蒸腾耗水与环境因子的耦合关系
在全球气候变化背景条件下,随着降雨的变化和夜间温度的升高,同时结合多个环境因素综合作用,使得对夜间液流的研究意义更大。由于观测技术的限制,目前对夜间液流的观测多集中在小尺度上(叶片、单株和冠层等),因此以往的研究大多只是在单个叶片或者单株尺度来分析夜间的蒸腾作用,而在较大尺度上的研究较少。虽然在叶片或单株尺度上植物蒸腾耗水可以忽略,但是研究尺度不同,研究结果存在很大差异,在样地或小流域尺度上,夜间的蒸腾耗水是生态水文模型的重要组成部分,如果将此忽略,可能造成模型模拟结果的不准确性,建议今后在建立蒸腾蒸散模型,可以反映出夜间蒸腾的部分,建立从叶片尺度的夜间蒸腾到个体和样地尺度的夜间蒸腾耗水与环境因子的耦合关系。
猜你喜欢
生态学报(2024年1期)2024-01-25 07:05:16
南京林业大学学报(自然科学版)(2022年1期)2022-11-29 10:10:16
生态学报(2021年6期)2021-04-25 08:24:42
农业工程学报(2021年3期)2021-04-15 10:03:14
节水灌溉(2020年9期)2020-09-16 00:32:12
农业机械学报(2020年3期)2020-04-01 08:11:10
种子(2019年5期)2019-07-02 11:37:44
电源技术(2017年1期)2017-03-20 13:38:00
天然产物研究与开发(2016年11期)2016-06-15 20:29:15
电源技术(2015年7期)2015-08-22 08:48:36
