
饥饿及恢复饲喂对牦牛生长性能、营养物质表观消化率和血清指标的影响
2018-12-13 06:01曾少玉王之盛彭全辉王立志邹华围祝伊枭周芯宇刘建忠
动物营养学报 2018年12期
曾少玉 王之盛* 彭全辉 薛 白 王立志 邹华围 胡 瑞 祝伊枭 周芯宇 刘建忠
(1.四川农业大学动物营养研究所,四川省牛低碳养殖与安全生产高校重点实验室,雅安625014;2.路德生物环保技术(古蔺)有限公司,泸州646509)
牦牛作为青藏高原地区的当家畜种,是当地人民赖以生存的生产和生活资料,为高原地区的经济发展做出了巨大贡献[1-2]。然而,牦牛的饲养方式还基本为传统的放牧饲养模式,这使得牦牛饲料来源和营养供给完全依赖于牧草的周期性枯荣变化。由于青藏高原独特的地理环境条件,在漫长的冷季容易出现大雪封冻的极端气候,冷季缺料使牦牛易于面临饥饿甚至断食的生存状态,“夏活、秋肥、冬瘦、春死”是青藏高原牦牛放牧饲养受自然条件约束的恶性循环现象。在经历1个冷季枯草期后,牦牛体重损失高达30%[2]。因此,饥饿是牦牛放牧养殖中的关键问题。饥饿是指动物在营养物质消化代谢后,能够并且想要进食,但是由于外界食物资源的限制,导致其无法进行采食的一种生物学状态[3]。饥饿状态下,动物为了满足机体代谢而利用葡萄糖(glucose,GLU),从而导致血糖水平降低[4]。为了维持血糖水平稳定,机体主要利用肝糖原分解及糖异生来生成血糖,从而为机体提供能量[5]。随着饥饿时间的延长,动物将最终耗尽体内存储的糖原[6]。随后,动物体将进行脂肪动员,体内的甘油三酯含量升高,并通过β氧化供能。而蛋白质则是动物最后的能量储备,如果饥饿时间过长,将会对动物造成严重损伤,甚至死亡[7-8]。研究发现,反刍动物经过营养限制后,恢复营养时会表现出一段快速生长时期,被称为补偿生长[9]。冷季能量有限使牦牛饥饿成为常态,而补偿生长在牦牛放牧系统中具有巨大的研究价值[10]。尽管已经有关于牦牛饥饿方面的研究[11-12],但牦牛在饥饿后恢复饲喂的相关资料还未见报道。本试验旨在比较牦牛正常饲喂期、饥饿期和恢复饲喂期3个阶段中,生长性能、营养物质表观消化率、血清代谢产物及相关激素含量的变化,阐明牦牛在饥饿期以及在恢复饲喂期间的适应情况,从而为提高牦牛面对恶劣环境的抗性研究积累数据并提供理论依据,为减少牦牛在冷季饥饿条件下体重损失,提高牦牛的生产性能提供数据参考。
1 材料与方法
1.1 试验动物
选取3周岁健康、体重相近的6头九龙牦牛[平均初始体重(183.60±15.62) kg]作为试验对象。
1.2 试验设计
参照Belanche等[13]的试验设计,将6头牦牛作为1个组。试验共56 d,其中预试期14 d,正试期42 d。正试期又分为正常饲喂期(normal feeding period,NFP,7 d)、饥饿期(starvation period,SP,7 d)以及恢复饲喂期(refeeding period,RFP,28 d),饥饿期牦牛断绝饲料,参照本课题组Yu等[11]以及任建林等[12]的研究对牦牛进行为期7 d的断食。
1.3 试验饲粮
在预试期、正常饲喂期和恢复饲喂期采用同一试验饲粮。试验饲粮配制借鉴我国《肉牛饲养标准》(NY/T 815—2004)中体重200 kg、日增重0.8 kg/d肉牛的营养需求,并参考本课题组Yu等[11]和任建林等[12]的配方,采用白酒糟+稻草+精料补充料方式,按照精粗比30∶70(干物质基础)进行混合后饲喂,其中精料补充料主要由玉米、麦麸、豆粕、菜籽粕、磷酸氢钙、碳酸钙、食盐、预混料等组成,粗料为稻草和白酒糟,试验饲粮组成及营养水平见表1。
1.4 饲养管理
试验在四川农业大学试验场进行,试验前对牦牛进行免疫、驱虫、消毒及编号等操作。试验期间对圈舍进行定期消毒。采用单栏单槽栓系饲喂。正常饲喂期及恢复饲喂期,每天09:00以及16:00进行饲喂,试验牛自由采食,根据预试期确定的采食量投喂饲粮,保证每次采食后槽内有10%左右的剩料。整个试验期间所有牛只自由饮水。
1.5 样品采集
在正常饲喂期及恢复饲喂期每周的第2~6天采用全收粪法进行消化试验[14]。从试验期第1天开始,每周第1天09:00前进行牦牛颈静脉采血,所采血液在阴凉处静置30 min以上,然后以1 006.2×g的离心力离心15 min,取血清,快速冷冻后-20 ℃保存待测[15]。
1.6 测定指标和方法
1.6.1 牦牛生长性能及营养物质表观消化率
记录每天的实际采食量,并换算成干物质采食量(dry matter intake,DMI),每次采血后对牦牛进行空腹称重,计算出每周的平均日增重(average daily gain,ADG),并计算出料重比(feed/gain,F/G)。参照AOAC(2002)[16]标准,测定饲粮及消化试验收集的粪便中水分、有机物(organic matter,OM)、粗蛋白质(crude protein,CP)、粗脂肪(ether extract,EE)、酸性洗涤纤维(acid detergent fiber,ADF)、钙(calcium,Ca)及磷(phosphorus,P)的含量。饲粮及消化试验收集的粪便中粗纤维(crude fiber,CF)含量参照GB/T 6434—2006中方法测定,中性洗涤纤维(neutral detergent fiber,NDF)含量参照Van Soest等[17]方法,采用Foss纤维仪(Fibertec 2010,丹麦)进行测定。

表1 试验饲粮组成及营养水平(风干基础)
1)预混料为每千克饲粮提供The premix provided the following per kg of the diet:Co (as cobaltous chloride 6-hydrate) 0.12 mg, Cu (as copper sulfate) 11.67 mg,I (as calcium iodate) 0.58 mg,Fe (as ferrous sulfate) 58.33 mg,Mn (as manganese sulfate) 23.33 mg,Se (as sodium selenite) 0.23 mg,Zn (as zinc sulfate) 35.00 mg,VA 30 000 IU,VD 12 000 IU,VE 90.00 IU。
2)综合净能为计算值[NEmf=DE×Kmf;Kmf=Km×Kf×1.5/(Kf+0.5×Km)。式中:NEmf为综合净能;Kmf为消化能转化为净能的效率;DE为饲粮的消化能;1.5为饲养水平值;Km为消化能转化为维持净能的效率;Kf为消化能转化为增重净能的效率。该计算方法参照我国《肉牛饲养标准》(NY/T 815—2004)],其余为实测值。NEmfwas a calculated value [NEmf=DE×Kmf; Kmf=Km×Kf×1.5/(Kf+0.5×Km. In the formulas, Kmfwas the conversion efficiency of DE to NE; DE was digestible energy of the diet; 1.5 was the value of feeding level; Kmwas the conversion efficiency of DE to net energy for maintenance; Kfwas the conversion efficiency of DE to net energy for gain. The computing method of NEmfreference to ChineseFeedingStandardofBeefCattle(NY/T 815—2004)], while the others were measured values.
营养物质表观消化率计算公式如下:
某营养物质表观消化率(%)=100×[食入的
该营养物质的量(g)-粪中该营养物质的量(g)]/
食入的该营养物质的量(g)。
1.6.2 血清生化指标
血清中GLU(试剂盒编号DRE-B1045c)、甘油三酯(trigluceride,TG)(试剂盒编号DRE-B6210c)、非酯化脂肪酸(non-esterified fatty acid,NEFA)(试剂盒编号DRE-B0793c)、总蛋白(total protein,TP)(试剂盒编号DRE-B6210c)、尿素氮(urea nitrogen,UN)(试剂盒编号DRE-B0544c)以及肌酸酐(creatinine,CRE)(试剂盒编号DRE-B0781c)的含量应用酶联免疫吸附测定(ELISA)试剂盒[购于卡迈舒(上海)生物科技有限公司]采用双抗体夹心法,使用酶标仪进行检测,具体操作步骤参考试剂盒所附说明。
1.6.3 血清激素指标
血清中胰岛素(insulin,INS)(试剂盒编号DRE-B6410c)、胰高血糖素(glucagon,GC)(试剂盒编号DRE-B6449c)、生长激素(growth hormone,GH)(试剂盒编号DRE-B6427c)、胰岛素样生长因子Ⅰ(insulin-like growth factor-Ⅰ,IGF-Ⅰ)(试剂盒编号DRE-B0732c)的含量应用ELISA试剂盒[购于卡迈舒(上海)生物科技有限公司]采用双抗体夹心法,使用酶标仪进行检测,具体操作步骤参考试剂盒所附说明。
1.7 数据统计与分析
数据用Excel 2010软件进行整理,采用SAS 8.1软件的Proc univariate过程进行数据正态性分布的检验,之后使用Proc GLM过程进行单因素方差分析,并采用Duncan氏法进行多重比较,试验数据以“平均值±标准差”表示,P<0.05为差异显著。使用Proc CORR过程进行指标相关性检验,P<0.05表示两者呈显著相关,相关系数的大小表明变量间相关性的强弱。
2 结果与分析
2.1 饥饿及恢复饲喂对牦牛生长性能的影响
由表2可知,牦牛在饥饿7 d后体重低于正常饲喂期,失重比例约达初始体重的10.06%(P>0.05)。恢复饲喂期前3周,DMI、F/G显著低于正常饲喂期(P<0.05);恢复饲喂期前2周ADG显著高于正常饲喂期(P<0.05);恢复饲喂期第4周,DMI、F/G以及ADG与正常饲喂期差异不显著(P>0.05)。

表2 饥饿及恢复饲喂对牦牛生长性能的影响
同行数据肩标相同或无字母表示差异不显著(P>0.05),不同字母表示差异显著(P<0.05)。下表同。
In the same row, values with the same or no letter superscripts mean no significant difference (P>0.05), while with different letter superscripts mean significant difference (P<0.05). The same as below.
2.2 饥饿后恢复饲喂对牦牛营养物质表观消化率的影响
由表3可知,饥饿后恢复饲喂的牦牛,在第1周时,CP、EE、CF、ADF以及P的表观消化率显著高于正常饲喂期(P<0.05),Ca的表观消化率则显著低于正常饲喂期(P<0.05);在第2周时,CP和P的表观消化率较正常饲喂期显著升高(P<0.05);在第3周时,Ca的表观消化率较正常饲喂期显著降低(P<0.05);而在第4周时,除了ADF的表观消化率显著高于正常饲喂期(P<0.05)和NDF的表观消化率显著低于正常饲喂期(P<0.05)外,其余营养物质表观消化率与正常饲喂期差异不显著(P>0.05)。DM和OM在整个恢复饲喂期均与正常饲喂期差异不显著(P>0.05)。

表3 饥饿后恢复饲喂对牦牛营养物质表观消化率的影响
2.3 饥饿及恢复饲喂对牦牛血清生化指标的影响
2.3.1 血清代谢产物含量的变化
由表4可知,在饥饿期,血清GLU、TG、UN和CRE含量较正常饲喂期显著降低(P<0.05),血清NEFA含量较正常饲喂期显著升高(P<0.05),血清TP含量则与正常饲喂期差异不显著(P>0.05)。而到了恢复饲喂期,在前2周,血清NEFA和TP含量显著高于正常饲喂期(P<0.05);在第1周和第3周,血清GLU含量显著低于正常饲喂期(P<0.05);在第2周和第3周,血清TG含量显著低于正常饲喂期(P<0.05);血清UN和CRE含量分别在第2周和第3周显著低于正常饲喂期(P<0.05);到了第4周,上述所有指标与正常饲喂期均无显著差异(P>0.05)。
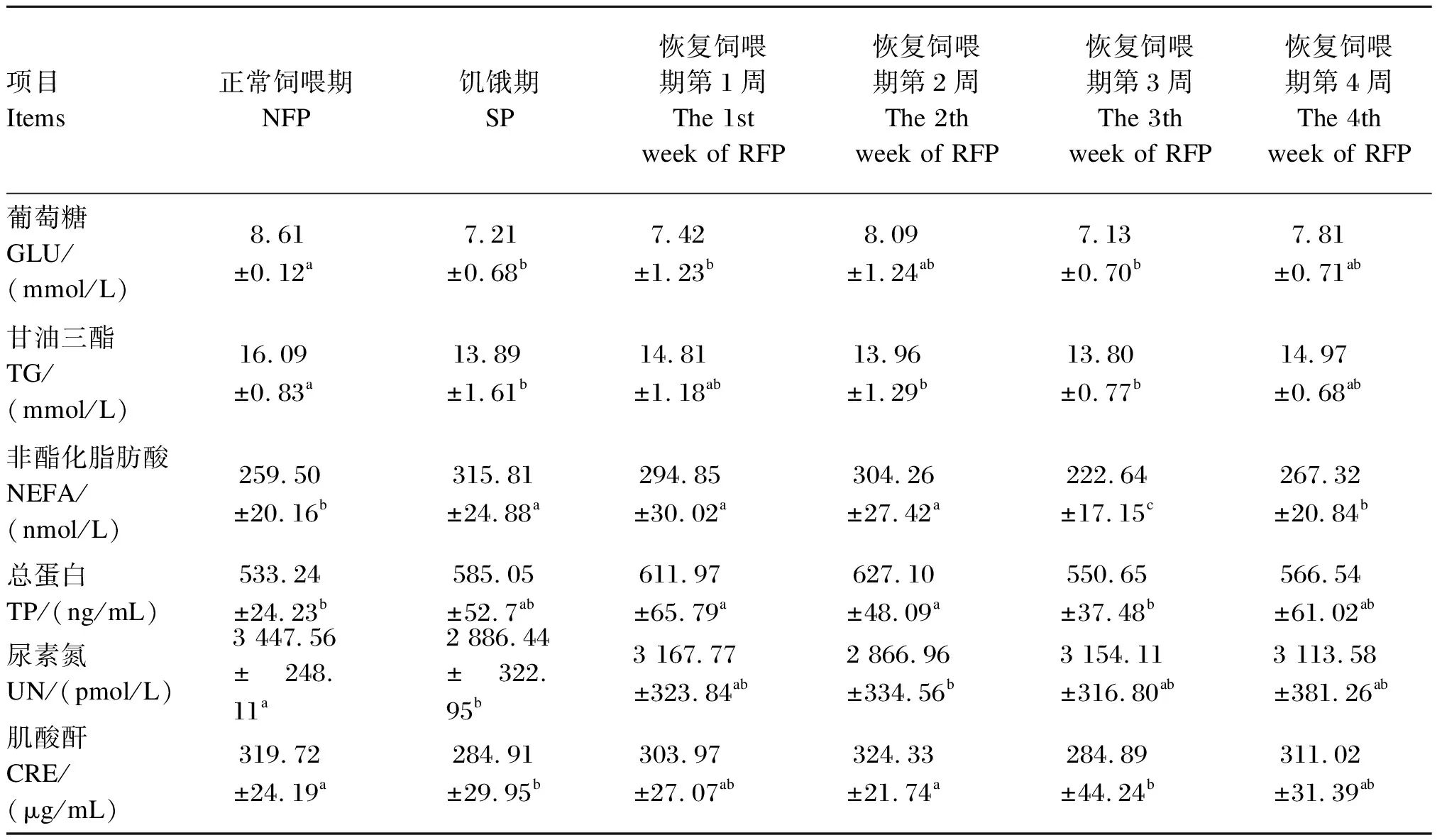
表4 饥饿及恢复饲喂对牦牛血清代谢产物含量的影响
2.3.2 血清激素含量的变化
由表5和表6可知,在饥饿期,血清INS、GH和IGF-Ⅰ含量及INS/GC较正常饲喂期显著降低(P<0.05)。而到了恢复饲喂期,在前3周,血清IGF-Ⅰ含量显著高于正常饲喂期(P<0.05);在第2周,血清INS含量显著高于正常饲喂期(P<0.05);在第4周,血清INS、GC含量及INS/GC显著低于正常饲喂期(P<0.05)。而血清GH和IGF-Ⅰ含量的相关性分析显示,在正常饲喂期为显著高度正相关关系(P<0.05),在饥饿期及恢复饲喂期第1周和第2周时无显著相关(P>0.05),在恢复饲喂期第3周呈现显著中度正相关关系(P<0.05),在恢复饲喂期第4周表现为显著中度负相关关系(P<0.05)。
3 讨 论
3.1 饥饿及恢复饲喂对牦牛生长性能的影响
在描述营养吸收后的动物无法摄取食物时,最常用的2个术语是饥饿和绝食[18]。饥饿主要是由于食物获取改变(如由食物资源充足变为资源匮乏)而造成的生理反应,而绝食则是由于内在机制调节下的生理反应,经常发生在动物躲避追捕、调节体温、脱毛及繁殖相关(如寻求配偶和领地防卫)的条件下[3]。饥饿研究中主要考虑2个因素,即饥饿时间和饥饿强度[19]。由于饥饿相关的研究尚未有一个统一的标准,考虑到牦牛冷季生长中饥饿时间和饥饿强度难以确定,在极端气候下甚至出现断水断料的情况,参考本课题组相关研究[11-12]后,对牦牛进行为期7 d的断食处理。
体重下降是动物饥饿后最显著的改变,饥饿期间OM的损失不可避免,而体重损失主要取决于动物的初始体重以及在饥饿期间各种能量资源的分配情况[20]。饥饿时动物体重减少的程度和速度可以用于衡量动物对饥饿的适应能力[21]。Chwalibog等[22]研究发现,猪在经过4 d低于维持饲粮水平25%的限制饲喂后,其体重损失达到初始体重的24%。Hiro-Omi等[23]发现鸡在经历6 d饥饿后,体重损失达到初始体重的35%。Chaiyabutr等[24]发现,山羊在经过2 d饥饿后,其体重损失就达到初始体重的16%。补偿生长是指动物在经历了一个限制饲料摄入的阶段后,恢复饲喂后具有更快生长潜力的一种生理过程[25]。研究表明,肝脏和胃肠道的大小改变与动物采食量的变化呈现等比例相关关系[26]。Ryan等[9]研究发现,在经过营养限制后,动物内脏器官变小并且生长减缓,而内脏器官重量的降低则促使机体降低维持能量需要,恢复饲喂后,动物内脏器官并不会立即恢复。肝脏和胃肠道对机体能量利用效率有着重要影响[27]。Connor等[28]发现,在恢复饲喂期间,饥饿后的肉牛ADG显著高于对照组肉牛,并且其细胞代谢、氧化磷酸化及三羧酸循环相关基因表达量显著升高。研究表明,恢复饲喂阶段,动物体重增长最快的是补偿生长的前期阶段[25,29]。在本次试验中,牦牛在经历7 d的绝食饥饿代谢后,体重损失为10.06%。与其他饥饿研究相比,牦牛饥饿强度和持续时间更高,但是体重损失更低,这说明牦牛对饥饿环境有较好的适应能力。恢复饲喂期前2周,ADG显著高于正常饲喂期,表现出明显的补偿生长现象。恢复饲喂期前3周DMI、F/G显著低于正常饲喂期,这可能是因为内脏器官变小,较小的肝脏和胃肠道使得机体能量利用效率升高,从而降低F/G。而到了恢复饲喂期第4周,ADG、DMI及F/G与正常饲喂期无显著差异,说明补偿生长主要在恢复饲喂期前3周。

表5 饥饿及恢复饲喂对牦牛血清激素含量的影响

表6 血清GH和IGF-Ⅰ含量的相关性变化
r>0表示两变量正相关,r<0表示两变量负相关。当|r|≥0.8时,认为两变量高度相关;当0.5≤|r|<0.8时,认为两变量中度相关;当0.3≤|r|<0.5时,认为两变量低相关;当0≤|r|<0.3时,认为两变量基本上不相关。P值表示两变量相关关系的显著性,其中P<0.05表示两变量相关关系显著,P>0.05则表示两变量相关关系不显著。
r>0 means positive correlation between two variables, andr<0 means negative correlation between two variables. When |r|≥0.8, it is considered that the two variables are highly correlated; when 0.5≤|r|<0.8, it is considered that the two variables are moderately correlated; when 0.3≤|r|<0.5, it is considered that the two variables are low correlated; when 0≤|r|<0.3, it is considered that the two variables are basically uncorrelated.P-value indicates the significance of correlation relationship between the two variables,P<0.05 means a significant correlation relationship between two variables, whileP>0.05 means no significant correlation relationship between two variables.
3.2 饥饿后恢复饲喂对牦牛营养物质表观消化率的影响
Jones等[30]发现,在进行营养限制后恢复饲喂,小鼠机体蛋白质合成和分解加快。Turgeon等[31]研究报道,羔羊在经过饥饿后恢复饲喂,出现了补偿生长现象,且在恢复饲喂早期阶段有较高的蛋白质沉积率。Lippens等[32]也发现,在经过营养限制后,恢复饲喂可以加快动物蛋白质周转,提高蛋白质利用效率。Heitz等[33]认为Ca消化率降低与P消化率的升高将会促使机体酸化作用加强,从而减少机体脂肪含量,加快蛋白质合成。Mehrez等[34]研究发现,NDF表观消化率降低与动物瘤胃内纤维降解菌和原虫数量的减少有关。Mahmoud等[35]也在小羊中的研究中得出相似结果。报道指出,NDF可以促进反刍动物咀嚼、唾液分泌以及瘤胃活动,确保瘤胃环境稳定以供瘤胃微生物的生长[36]。本研究中,整个恢复饲喂期DM和OM表观消化率与正常饲喂期差异不显著,与Ma等[37]研究结果相似。恢复饲喂期第1周,牦牛对饲粮中CP及EE的表观消化率升高,同时Ca和P表观消化率在第1周和第2周时也发生了变化,有利于脂肪分解和蛋白质合成,从而促进补偿生长的发生。恢复饲喂期第4周,除ADF和NDF表观消化率外,其他营养物质表观消化率与正常饲喂期差异不显著。NDF的表观消化率在恢复饲喂期第2周、第3周和第4周显著降低,这种变化可能与瘤胃环境及瘤胃微生物改变有关。
3.3 饥饿及恢复饲喂对牦牛血清代谢物含量的影响
动物机体所有组织都利用GLU,GLU是中枢神经系统、肾髓质以及成熟红细胞首选的能量物质,必须保证其持续供应,饥饿条件下,外源能量不足,导致机体不断消耗自身的GLU,最终使得血糖水平降低[38]。动物在饥饿状态下主要通过分解肝糖原来生成GLU供能,同时,利用糖异生作用维持血糖水平稳衡[5]。在肝脏和肾脏中,GLU可以由氨基酸(主要是丙氨酸)、甘油(主要来源于TG的水解)、酮体以及循环的乳酸和丙酮酸进行从头合成[39]。而到了恢复饲喂阶段,随着食物的摄入,血清中GLU含量升高,INS分泌量升高,而INS又促进动物吸收GLU并转运到特定的细胞中利用,最终导致血糖水平降低[40]。本研究中,饥饿期血清GLU含量较正常饲喂期显著降低,这可能是机体为了适应饥饿而降低糖代谢水平[12]。恢复饲喂期,血清GLU含量在第1周和第3周显著降低,这可能是因为动物在补偿生长阶段各细胞组织对能量的需求较高,促使机体对糖的利用较高。
在低血糖的条件下,脂肪酸是机体能量需求的一种可行性来源[41]。Blum等[41]发现,在营养限制时期,受到限制的牛血清中非酯化脂肪酸的含量显著高于非限制的牛只组,这体现出较强的脂肪动员。饥饿期间脂肪动员提高是为了确保机体重要蛋白质含量的同时为机体提供足够的能量[7]。Dimarco等[42]研究发现,在恢复饲喂阶段,受到饥饿限制的牛只,血清NEFA含量在前8天的时间都保持较高的水平,说明在恢复饲喂开始时,动物的脂肪代谢并没有恢复到正常饲喂期水平。本研究中,饥饿期血清TG和NEFA含量的变化体现了较强的脂肪动员。在恢复饲喂期前3周,血清NEFA含量在前2周显著高于正常饲喂期,在第3周则显著低于正常饲喂期,而血清TG含量在第2周和第3周显著降低,说明在前3周牦牛代谢状态还未恢复到正常时期水平,还保持着较高的脂肪分解水平。
血清UN含量是衡量蛋白质周转代谢的重要指标,UN含量升高说明蛋白质周转代谢加快[29]。CRE是磷酸肌酸的分解产物以及肌肉组织肌酸代谢的终产物,血清CRE含量的降低说明肌肉蛋白质分解代谢减慢[43]。本研究中,血清UN含量在饥饿期显著降低,这与Harlow等[44]在草原土拨鼠上的研究结果以及Costa等[45]在象海豹上的研究结果相似,说明在饥饿期间蛋白质周转降低,而血清CRE含量降低说明饥饿期间肌肉组织蛋白质分解减慢。由此可知,机体在饥饿期间通过降低蛋白质的代谢水平来减少蛋白质消耗。恢复饲喂期间,前2周血清TP含量显著升高,这表明在恢复饲喂期前2周蛋白质沉积增加,这有利于牦牛在恢复饲喂后出现补偿生长现象。
3.4 饥饿及恢复饲喂对牦牛血清激素含量的影响
INS促进靶细胞对血液中GLU的摄取和转运,从而降低血糖水平[40]。而GC则促进脂肪组织的分解,提高血糖水平,对INS有拮抗作用,血糖水平的升高会使得INS/GC降低,进而提高血糖水平[46]。在许多饥饿代谢的研究中都观测到动物血液中INS含量显著下降的现象,饥饿期间血液中INS含量的降低和GC含量的升高有利于机体利用脂肪,维持血糖水平[47]。Ahmed等[48]报道,受到营养限制的牛只在之后的恢复饲喂期间体内INS的含量快速升高,这种变化与氨基酸转运加快、氧化减慢以及蛋白质降解减慢有关。Blum等[41]也发现了相同的变化趋势,在恢复饲喂期间,胰岛素分泌量的增加启动了体内合成过程的信号,动物合成代谢增强。Shaw等[49]报道,INS是促进机体肌内脂肪合成最重要的激素。本研究中,饥饿期间,牦牛血清INS含量和INS/GC显著降低,这有利于机体利用脂肪分解提供能量。恢复饲喂后,牦牛血清INS含量在第2周显著升高,这有利于促进机体的合成代谢,从而促使牦牛出现补偿生长现象。
动物在进食后,GH的分泌会促使肝脏合成IGF-Ⅰ,而IGF-Ⅰ通过分配各种营养成分来促进组织及机体生长[50]。研究发现,动物在受到食物限制后,GH和IGF-Ⅰ含量的相关关系变得复杂[51]。Keogh等[52]认为,GH和IGF-Ⅰ含量相关性改变可能是由于营养限制使得肝脏变小的原因。Li等[53]报道,在能量限制期间,动物机体内GH及其高亲和力的受体含量会显著降低,同时两者在肝脏内的特异性结合能力也会下降,而IGF-Ⅰ含量的降低慢于生长激素受体含量的降低。有研究发现,受到营养限制的试验动物机体IGF-Ⅰ含量显著降低,但是GH含量的变化却不显著[54]。IGF-Ⅰ含量的降低可能是由于机体对营养限制的一种适应手段,将细胞的生长调节到最慢,从而将有限的能量运用到重要的细胞功能上面,比如组织的维持以及修复[53]。Bell等[55]推测,这种代谢的改变可能是为了保证动物在营养限制期间体内代谢的稳态。本研究中,饥饿期间,血清GH和IGF-Ⅰ含量显著降低以及两者相关关系的变化,有利于减缓细胞生长,节约能量,保持代谢稳定,从而适应饥饿环境。恢复饲喂后,血清GH含量恢复正常饲喂期水平,血清IGF-Ⅰ含量在前3周显著升高,两者相关性在恢复饲喂期发生变化,可能是因为肝脏尚未恢复导致两者之间代谢关系维持在饥饿期的原因,而这种代谢状态的维持则有利于补偿生长的产生。
4 结 论
① 牦牛在经历7 d的饥饿后,表现出对营养限制较好的适应现象。牦牛在饥饿期间通过降低糖代谢和蛋白质周转水平以及提高脂肪分解代谢来适应饥饿。
② 从牦牛ADG、F/G及血清生化指标进行评价,可以得出,牦牛在饥饿后恢复饲喂,补偿生长主要在前3周,到了第4周即与正常饲喂期无显著差异。
猜你喜欢
河北果树(2021年4期)2021-12-02
金桥(2021年10期)2021-11-05
上海公路(2019年3期)2019-11-25
湖南饲料(2019年5期)2019-10-15
福建基础教育研究(2019年10期)2019-05-28
特产研究(2016年3期)2016-04-12
文学少年(原创儿童文学)(2016年19期)2016-02-28
文史春秋(2016年8期)2016-02-28
动物营养学报(2015年10期)2015-12-01
小说月刊(2014年8期)2014-04-19
