
暖季型水生植物残体分解对冬季浮床氮去除效果的影响
2018-12-05 06:47吕丽萍张淼陈琛安树青赵德华
生态环境学报 2018年11期
吕丽萍,张淼,陈琛,安树青,赵德华
南京大学生命科学学院,江苏 南京 210023
水生植物是人工湿地的重要组成部分,通过直接吸收、根系泌氧、有机质分泌和为微生物提供附着基质等途径,对人工湿地的净化功能产生重要影响;同时,水生植物残体在分解过程中也会释放富营养物质,对人工湿地造成二次污染。因此,对于水生植物在人工湿地中的功能一直存在争议(Brisson et al.,2009;Liu et al.,2014;Cardinal et al.,2014)。长江中下游地区,人工湿地工程中多种植芦苇(Phragmites australis)、菖蒲(Acorus calamus)等暖季型水生植物,而暖季型水生植物残体在秋、冬季节的集中腐解,极易对人工湿地造成二次污染,因此在人工湿地管理中,常通过收割移除水生植物残体,降低二次污染的风险(刘臣,2014)。
随着尾水湿地深度净化工程技术的广泛应用,尾水湿地的ω(C)/ω(N)失衡问题越来越受到研究者的重视,外加植物碳源在缓解尾水湿地ω(C)/ω(N)失衡、提升氮去除效率方面表现出了极大的潜力(Hang et al.,2016;Fu et al.,2016),协调尾水湿地中植物碳源添加与水生植物残体收割移除,以及水生植物的净化功能与二次污染风险间的矛盾,已成为尾水湿地工程应用中的重要内容(Chen et al.,2014a;Zhang et al.,2016)。在尾水湿地中通过将不同物候型水生植物进行合理配置可以协调这一矛盾,且能够达到稳定周年出水水质的目的(张惠等,2017)。关于水生植物残体分解过程及其对水环境的影响(Kayranli et al.,2010)、不同物候型水生植物净化功能提升及其机制等方面已有大量的研究报道(Wu et al.,2017),并初步探讨了不同物候型水生植物配置提升人工湿地净化功能的可行性(Zhang et al.,2016;张惠等,2017)。目前对于暖季型水生植物残体在枯萎期的集中分解与冷季型水生植物在生长期的净化功能间的相互作用研究少见报道,而此类研究对于完善不同物候型水生植物配置理论及其工程应用具有重要的意义。
本研究以长江中下游地区广泛采用的人工湿地植物——芦苇为材料,研究芦苇残体在冬季分解过程中对冷季型水生植物——水芹菜(Oenanthe javanica)的生长,以及对水芹菜浮床氮去除率的影响,旨在提出一种同步提升尾水湿地氮去除功能和降低水生植物二次污染风险的解决方案,并为其工程应用提供理论依据。
1 材料与方法
1.1 试验设计
试验在南京大学洪泽湿地实验基地(33°19'N,118°55'E)的塑料大棚中进行,试验基地所在流域年降水量1080 mm,年平均温度为15 ℃,冬季平均温度为2~5 ℃。
试验选取人工湿地中常见的水生植物芦苇作为碳源添加物质。在枯萎期(12月21日)收集基地附近池塘的芦苇残体,将收集的芦苇残体洗净、晾干,剪成2~3 cm的小段,在70 ℃烘箱烘干至恒重,分装在尼龙袋(43 cm×34 cm,网眼2 mm)里储存于干燥箱中。选用冬季植物水芹菜为浮床植物,水芹菜从当地的苗圃中收集,选用株高7~8 cm且长势一致的幼苗,将它们的根切成相同的长度(2 cm)。然后将这些幼苗清洗干净,待幼苗制备完毕后移植到浮床上。
试验装置为长190 cm、宽110 cm、高66 cm的纤维增强塑料长桶,设置2个处理组:不加芦苇秸秆浮床系统(CK)和添加芦苇秸秆浮床系统(FCW-L),其中,FCW-L的芦苇秸秆添加量为每个系统中加10 kg芦苇(即5 kg·m-2,由于实际湿地中芦苇的生物量变化较大,最佳用量较难确定,本研究取中间值)。每处理组设置3个重复,共6个浮床人工湿地系统单元。试验始于2018年1月15日,结束于2018年4月5日。以HCFC500型生态浮床(500 mm×500 mm×65 mm,每个浮床有9个栽植篮,孔径8 cm)作为水芹菜的生长载体,每个湿地系统6块浮床共54篮浮床植物,每个水培篮中移植4株幼苗,浮床面积占整个湿地单元表面积的72%。每个系统每篮水芹菜的初始生长状况和生物量基本一致,各单元水位设定为40 cm。
1.2 试验运行
试验用水取自基地附近池塘,水质为:化学需氧量(COD)35 mg·L-1,铵态氮(NH4+-N)1 mg·L-1,硝态氮(NO3--N)0.5 mg·L-1。以池塘水为基础,添加由 KNO3和(NH4)2SO4配制而成的人工合成污水,以20 d为1个周期,每个周期NH4+-N添加量为 10 mg·L-1,NO3--N 添加量为 5 mg·L-1(每周期添加1次,上一周期剩余的营养物质累加到下一周期)。系统共运行4个周期(80 d),每个周期结束时取水样和植物样品,测定水质及植物指标。
在整个试验期间,近地层大气温度波动范围为-0.11~25.2 ℃,平均温度为 10.5 ℃,而水温波动范围在-1~26.1 ℃之间,平均温度为10.7 ℃,温度逐渐升高(图 1)。各周期的平均气温温度分别为5.23、7.48、10.21和16.19 ℃,各个周期平均水温分别为6.03、7.05、10.25和16.3 ℃。
1.3 样品采集与测定
在每个试验周期结束时,于水下20 cm处采集水样,利用哈希 DR/2800分光光度计对 COD、NH4+-N、NO3--N和总氮(TN)进行测定,COD浓度测定采用USEPA消解比色法,NH4+-N浓度测定采用水杨酸法,NO3--N浓度测定采用镉还原法,TN浓度测定采用过硫酸盐氧化法(张惠,2017);水体溶解氧(DO)浓度及 pH测定使用美国哈希HQ40d双路输入多参数数字分析仪;使用 HOBO防水型温度光照记录仪(UA-002-64)监测系统水温及气温变化。同时每个系统取6篮水芹菜用于植物指标测定,其中3篮用于测量其株高、干重及有机碳(C)和总氮(N)含量。在70 ℃烘箱中烘至恒重,称量干重,C含量测定采用重铬酸钾容量法-外加热法(NY/T 1121.6—2006),N含量测定采用凯氏定氮法(鲍士旦,2000)。另外 3篮水芹菜用于测定叶绿素和根系活力,叶绿素使用 SPAD502叶绿素含量测定仪进行测定(邹祥旭,2016),根系活力采用TTC 法测定(白宝璋等,1994)。每次取完样品后及时补充水芹菜以保证整个试验周期植物种植密度不变。在第5天、20天、80天(考虑到芦苇前期分解较快,生物量及成分变化较大,后期分解缓慢,且频繁取样对试验结果影响较大,故在前期增加对芦苇的测样(5 d),后期40 d和60 d不再对芦苇进行取样,以减少实验误差。),从各装置中取1袋芦苇,在70 ℃烘箱中烘干至恒重测定其生物量及成分变化,测定方法与水芹菜相同。
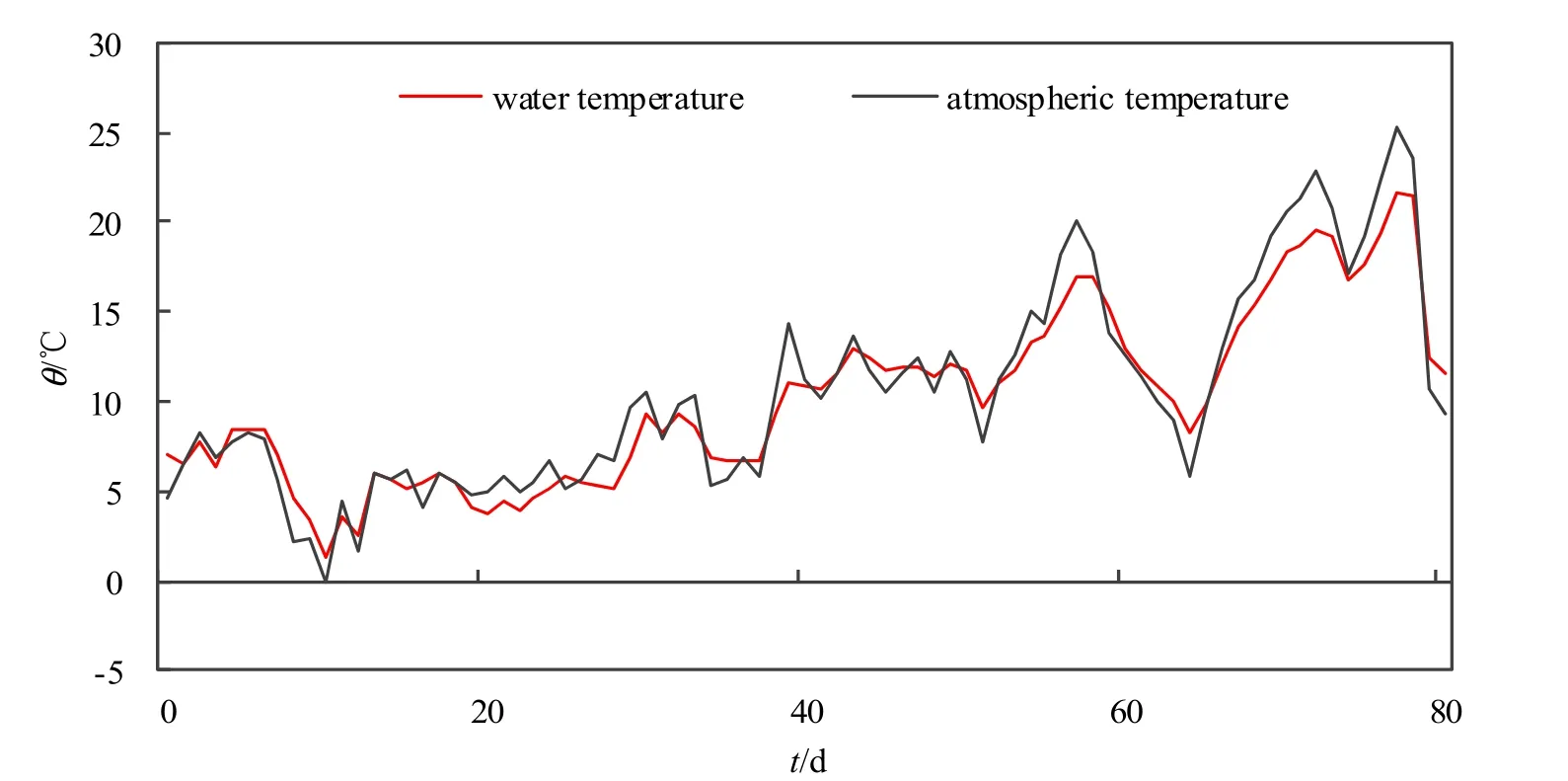
图1 试验期间水温和气温变化Fig. 1 Changes of water and air temperature over time
2 结果与分析
2.1 芦苇分解过程中的质量与成分变化动态
芦苇残体分解表现出先快后慢的动态过程(图2),试验前5天,芦苇残体质量减少了18.39%,经过80 d的分解,质量累计减少33.4%。图3所示为芦苇残体中C、N含量及ω(C)/ω(N)的动态变化,C含量在整个试验过程中呈下降趋势,在前 20天 C含量下降较快,由最初的49.02%下降至41.87%,随后下降缓慢,20~80 d C含量下降4.51%。N含量在前20天从0.74%下降至0.33%,后60天N含量有所上升,在第80天时升高到0.42%。ω(C)/ω(N)表现为先升高后降低的趋势,在第 60天达到最大值(128),后20天从128下降到89.6。芦苇残体的这种先快后慢分解动态是水生植物分解的典型过程。由此表明,对于C缺乏的人工湿地系统,芦苇在分解前期可向系统中释放大量C源以满足反硝化脱氮的需要。
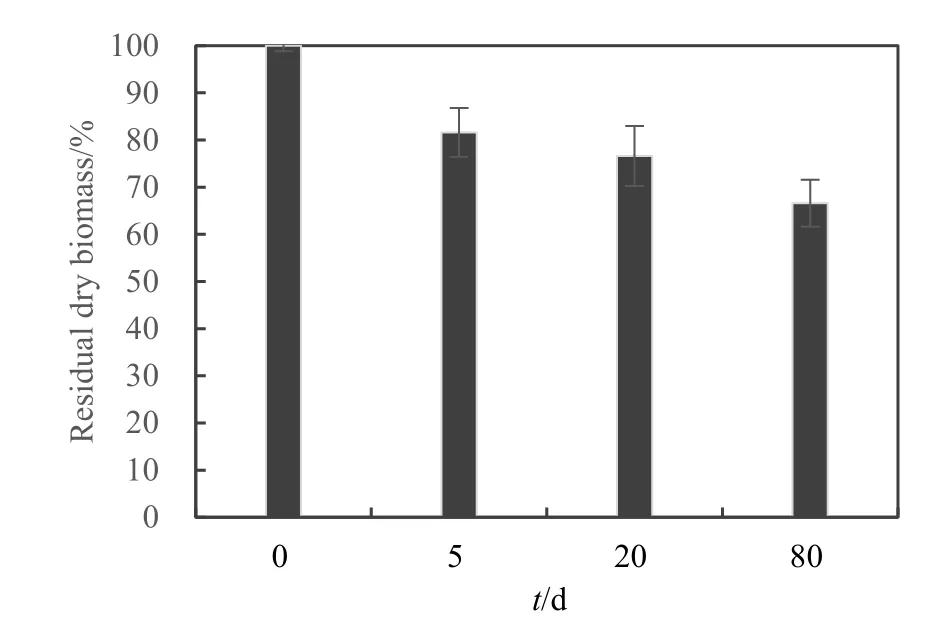
图2 芦苇分解过程中剩余质量的变化Fig. 2 Temporal dynamic of residual dry biomass of Phragmites communis
2.2 芦苇分解对水芹菜生长的影响
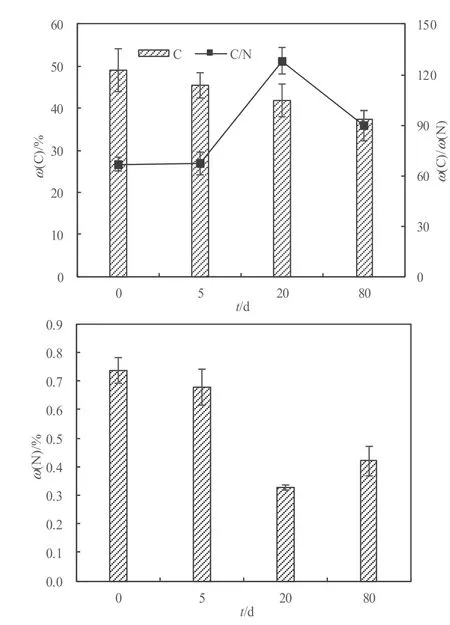
图3 芦苇分解过程中C含量、N含量及ω(C)/ω(N)动态变化Fig. 3 Temporal dynamics of C concentration, N concentration and ω(C)/ω(N)in Phragmites australis residue
试验期间,两个处理组的水芹菜根长和株高在前两个试验周期均增长缓慢(图 4);后两个周期FCW-L水芹菜根长和株高增长迅速,根长由第2周期结束时的7.07 cm增加至第4周期结束时的12.53 cm,而株高从10.03 cm增长至33.57 cm;CK在第3周期生长较快,根长由6.33 cm增加至9.37 cm,株高由8.37 cm增加至14.07 cm,第4周期,根长和株高分别缩减至4.70 cm和12.03 cm。生物量与株高表现为相似的趋势,FCW-L干重从第3周期开始迅速增加,地上干重由第 2周期结束时的 2.67 g·m-2增至第3、4周期的33.75 g·m-2和101.73 g·m-2,地下干重由 4.26 g·m-2增至 13.47 g·m-2和 20.76 g·m-2;CK 的地上干重由初期的 2.91 g·m-2增至第 3周期的 8.88 g·m-2,第 4 周期降至 3.78 g·m-2;地下干重由初期的 1.24 g·m-2增至第 3周期的 3.21 g·m-2,第 4 周期降至 0.51 g·m-2。由此表明,芦苇残体的分解可促进水芹菜的生长,从而提升冬季浮床的净化功能。
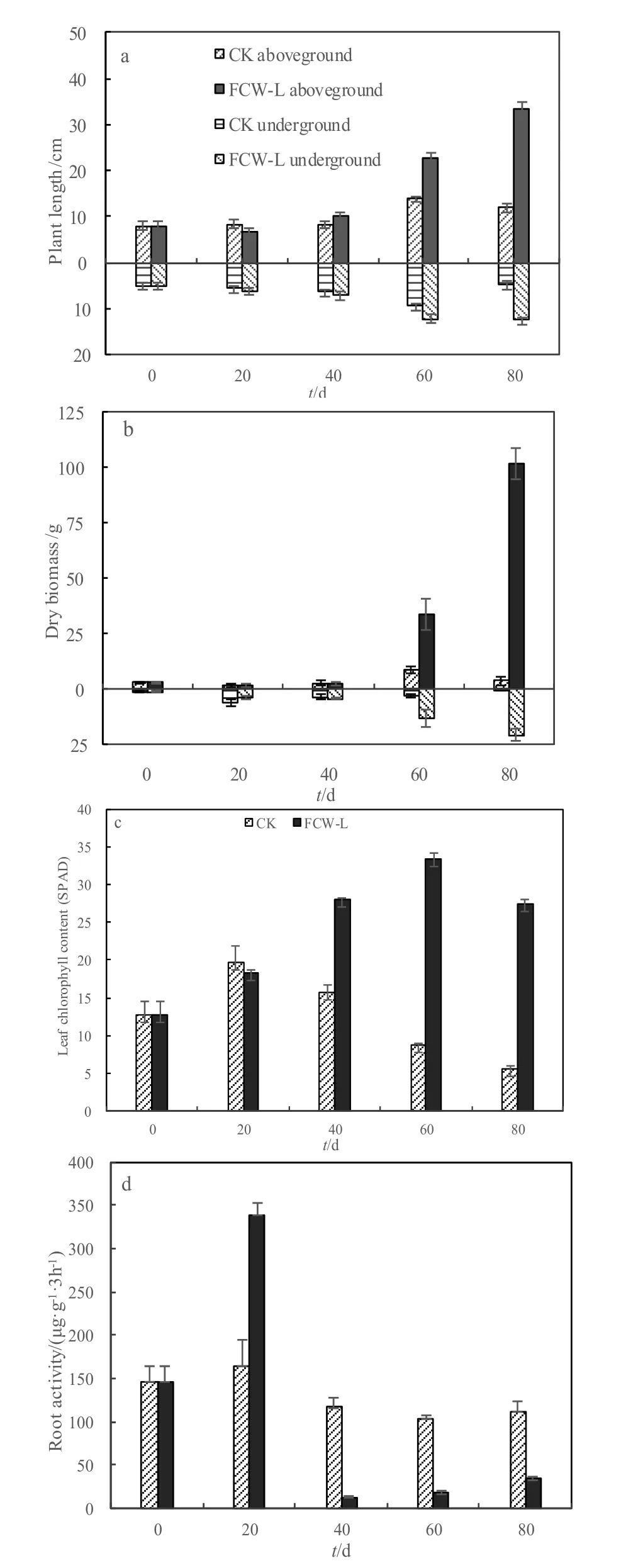
图4 浮床植物水芹菜的生长动态:株高(a)、干重(b)、叶绿素(c)、单位根系活力(d)Fig. 4 Growth characteristics of plant length (a), dry biomass (b), leaf chlorophyll concentration (c) and root activity (d) of the Oenanthe clecumbens plants
FCW-L的叶绿素含量表现为前3个周期持续增长,第4周期略有下降;而CK从第2周期开始表现为持续下降趋势。两处理间叶绿素在第1周期无显著差异(P=0.392),但从第2周期到第4周期,FCW-L叶绿素含量显著高于 CK(P=0.000)。CK的单位根系活力在第一周期略有上升,随后3个周期保持在 100~120 μg·g-1·3h-1,而 FCW-L 的水芹菜单位根系活力在第 1周期结束时达到最大值(339 μg·g-1·3h-1),第 2 周期迅速下降,随后基本稳定在10~30 μg·g-1·3h-1;总根系活力 FCW-L 在各周期显著高于CK(P=0.000~0.001)。结果表明,添加芦苇残体可提高水芹菜叶绿素含量和总根系活力,而降低单位根系活力。
2.3 芦苇分解对水芹菜浮床净化功能的影响
2.3.1 pH、电导率和DO
试验期间,两个系统总体上处于弱碱性,FCW-L组和CK的pH表现出先上升后下降的趋势,pH在第一周期升高,20 d时分别达到最大值9.94、8.66,后3个周期pH下降(图5)。两个处理组的电导率一直呈上升趋势,在第4周期结束时分别达到最大值 1784.70 μs·cm-1、1779.00 μs·cm-1。前 3个周期 FCW-L系统的 pH值显著低于 CK(P=0.000~0.017),FCW-L电导率显著高于 CK 组(P=0.000~0.012),而第4周期两处理组的pH和电导率均没有显著差异(P=0.213,0.7)。各周期,FCW-L处理DO均显著低于CK(P=0.000),FCW-L一直处于厌氧状态,而CK一直处于好氧状态;CK的DO从前两个周期的接近于饱和状态降至第3、4周期的 3.97 mg·L-1和 4.84 mg·L-1。FCW-L 组在加入芦苇后溶氧迅速下降,在第4天达到最低值(0.18 mg·L-1),随后一直处于缺氧环境(DO波动范围为0.18~0.7 mg·L-1)。由此表明,芦苇残体分解会引起系统内DO显著下降,使系统处于厌氧状态,同时对部分周期的pH和电导率也产生显著的影响。
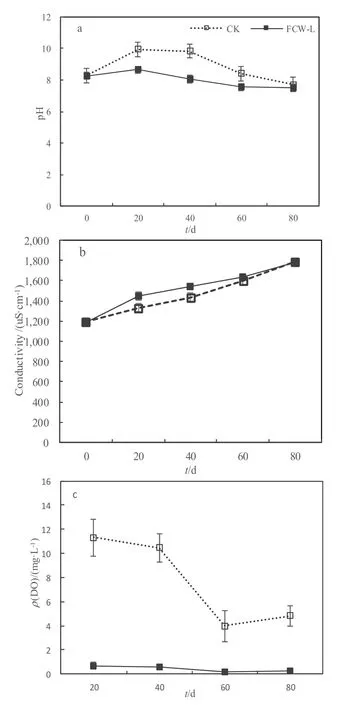
图5 浮床系统中pH(a)、电导率(b)和溶氧(c)变化Fig. 5 Field measurements of pH (a), conductivity (b) and DO (c)in the FCW
2.3.2 NO3--N、NH4+-N和TN
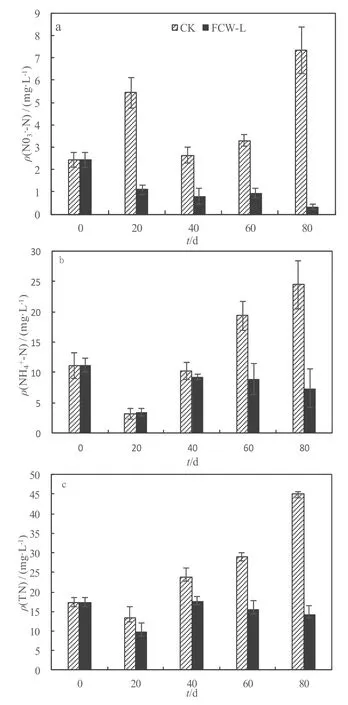
图6 硝氮(a)、氨氮(b)和总氮(c)质量浓度变化Fig. 6 NO3--N (a), NH4+-N (b) and TN profile (c) in CK and FCW-L
由图6可知,各试验周期结束时,CK的NO3--N浓度均显著高于 FCW-L(P=0.000~0.003),其中CK的NO3--N波动范围为2.63~7.45 mg·L-1,均值为 4.68 mg·L-1;FCW-L 的 NO3--N 波动范围为0.33~1.1 mg·L-1,均值为 0.79 mg·L-1,4 个周期表现为逐渐下降的趋势。FCW-L 4个周期NO3-N的累计去除率为90%,而CK 4个周期NO3--N的累计去除率仅为26.5%。NH4+-N在前两个周期结束时处理间无显著差异(P=0.651,0.316),后两个周期结束时 FCW-L的 NH4+-N显著低于 CK(P=0.007,0.005)。CK各个周期末NH4+-N表现为逐渐上升的趋势,各周期末的波动范围为3.47~24.43 mg·L-1,均值为14.31 mg·L-1;FCW-L第2周期末与第1周期末相比显著上升(P=0.009),而后3个周期结束时差异不显著(P>0.05),各个周期末波动范围为3.47~9.27 mg·L-1,均值为 7.27 mg·L-1。4 个周期末FCW-L和CK的NH4+-N累计去除率分别为82%和41%。TN变化趋势与NH4+-N相似,CK的TN呈上升趋势,FCW-L在第二周期结束达到最高随后开始下降,整个周期CK和FCW-L的TN去除率分别为27.2%和73.4%。由此表明,芦苇残体的分解可提升水芹菜浮床湿地NO3--N、NH4+-N和TN的去除率。
2.3.3 COD
由图7可知,各个周期结束时,FCW-L的COD均显著高于CK(P=0.000),其中,FCW-L在各个周期末的波动范围为 171.33~196.67 mg·L-1,平均值为184.67 mg·L-1;而CK在各个周期末的波动范围为 56.00~78.33 mg·L-1,平均值为 72.08 mg·L-1。FCW-L 各个周期末的ω(C)/ω(N)[ω(COD)/ω(TN)]均显著高于CK(P<0.05),FCW-L和CK各个周期末ω(C)/ω(N)的 波 动范围分别为 1.73~5.82 和9.72~18.07,平均值分别为3.18和13.10。由此表明,添加芦苇残体可提升系统有机物含量,改善碳氮失衡状况。
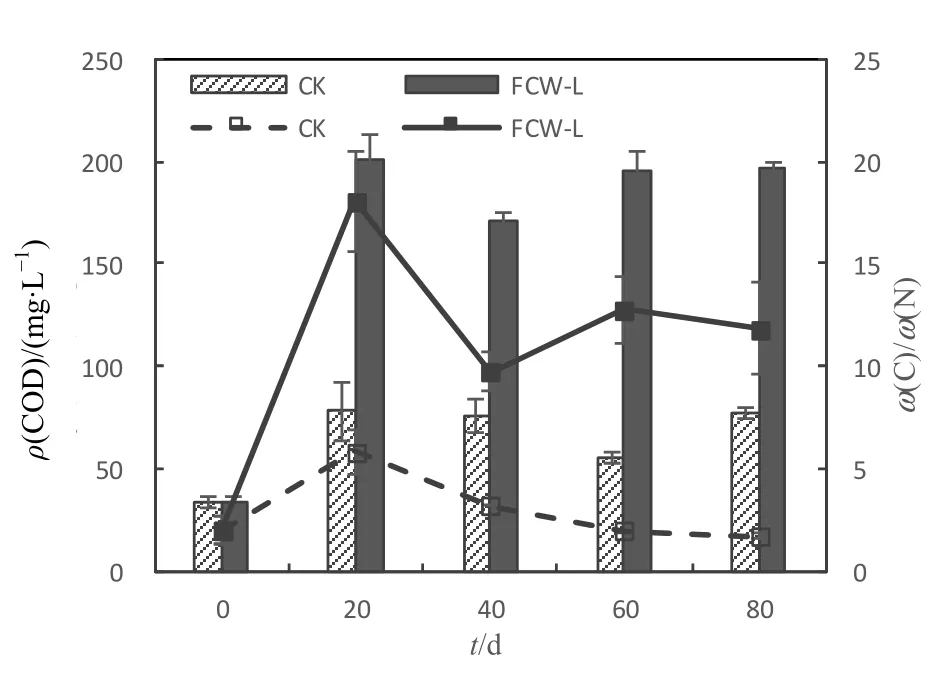
图7 COD质量浓度和碳氮比变化Fig.7 COD and ω(C)/ω(N) profile in CK and FCW-L
3 讨论
3.1 冷季型水生植物生长的影响因素及其与植物残体分解的关系
温度是影响植物生长的重要因素,温度通过影响植物的光合作用、呼吸作用、细胞分裂和伸长来影响植物的生长(袁伟,2006)。水芹菜生长的最佳温度为12~24 ℃,高于25 ℃或低于10 ℃均不利于其生长,温度为 10 ℃时水芹菜会出现不正常的外部特征,下部叶片开始发黄,茎细且植株生长缓慢(胡绵好等,2011)。试验前两个周期气温低于 10 ℃,水芹菜生长缓慢,叶绿素和根系活力相对较低,后期随着温度回升,FCW-L的水芹菜生长迅速,株高和生物量增加。溶氧是水环境中重要的生态因子,对植物的生长存在影响(许建平等,2014)。FCW-L的水芹菜由于系统DO降低,后3个周期的根系活力较低,这与李宽意等(2006)的研究结果一致。在低氧条件下,植物为了生存会在根茎基部产生不定根,替代死亡或受到伤害的初生根吸收水分和养分以维持植物生长(Sauter,2013)。尽管 FCW-L的芦苇秸秆分解形成低氧环境对植物生长造成影响,水芹菜单位根系活力下降,但低氧胁迫促进了水芹菜不定根的生长,根系生物量大大增加。CK的水芹菜生长缓慢,到第4周期甚至出现枝叶枯萎和根系腐烂,这可能是由于浮床人工湿地中缺少植物生长所必须的微量元素,而 FCW-L的芦苇秸秆在微生物作用下分解可以提供微量元素(杨芳,2011)。劳秀荣等(2002)研究表明秸秆还田量的增加使土壤有效锌含量明显提高。于淼等(2015)研究也发现秸秆生物降解明显增加了土壤有效铜的含量,提高了红干椒(Capsicum annuum)植株中的铜元素含量。易文利等(2011)研究表明,植株萌发的初期,投加一定量的有机质(植物残体),短时间内穗花狐尾藻(Myriophyllum spicatum)根系由于厌氧环境而受到一定抑制,但很快恢复生长,且中后期可利用的营养盐质量浓度较低,又促进根系生长,从而促进了植 株的生长。FCW-L的芦苇残体分解对水芹菜生长没有明显的抑制作用,且水芹菜根系生物量大大增加,总根系活力提高。
3.2 植物残体分解提升浮床湿地冬季氮去除效果及其可能途径
人工湿地中存在多种脱氮机制,包括植物吸收、基质吸附、硝化反硝化等,其中,通过微生物代谢过程进行硝化-反硝化作用是人工湿地脱氮的主要途径(Changgyun et al.,2009)。反硝化过程需要有机碳作为电子供体,以还原硝酸盐氮。因此,有机碳源是反硝化过程中必不可少的物质(刘刚等,2011)。Wen et al.(2010)研究表明,硝氮去除率与可有效利用的碳源呈强相关(R2=0.93)。Chen et al.(2014a)研究也表明,添加香蒲(Typha orientalis Presl)凋落物增加了人工湿地系统的碳源,硝氮去除率增加了 1.2~1.3倍。在本研究中,添加芦苇秸秆大大改善了系统中碳源缺乏的问题,FCW-L的ω(C)/ω(N)远高于CK,硝氮去除率也相应增加。Fan et al.(2013a)研究,表明进水ω(C)/ω(N)从 5 增加到10,由于碳源增加人工湿地的出水NO3--N浓度从 23 mg·L-1降低到 4.72 mg·L-1,FCW-L 芦苇腐解提供足够的碳源促使硝氮去除率大大提高。同时,在人工湿地中添加芦苇秸秆,其分解过程会中消耗大量氧气,有利于创造反硝化细菌所需要的缺氧环境(Chen et al.,2014a)。在试验期间,FCW-L一直处于缺氧状态,有利于反硝化作用的进行,硝氮净化效果提升。有研究表明,以植物作为外加碳源的人工湿地,对硝氮的去除效果会随着时间的延长而降低(Chen et al.,2014a;Chen et al.,2014b)。但本试验结果与之相反,FCW-L 4个周期的硝氮去除率呈波动上升,这可能是由于植物种类不同。本试验中的芦苇为挺水植物,木质素等难降解物质含量较高,与浮叶植物和沉水植物相比,挺水植物分解周期长(Kayranli et al.,2010)。因此,芦苇作为反硝化的植物碳源后期不会出现由于碳源不足而导致硝氮去除率降低。此外,温度也是影响硝氮去除效果的重要因素,微生物活性与温度密切相关(Zhang et al.,2016),试验后期温度升高有利于浮床人工湿地微生物活动,微生物途径去除硝氮作用也会随之增强。
溶氧和植物是影响氮去除的重要因素(Moss,1990)。因为硝化过程需要氧,所以在硝化动力学中DO是一个极其重要的因素(张政等,2006)。FCW-L在芦苇添加后一直处于无氧状态(DO<1 mg·L-1),而溶氧高于 1.5 mg·L-1是硝化作用发生的条件(Ye et al.,2009),因此理论上FCW-L不能满足硝化作用去除氨氮的条件。同时CK中的溶氧初期接近饱和溶氧,后期溶氧降低但仍然满足硝化作用发生的先决条件。本试验中,FCW-L 4个周期的氨氮去除率均高于CK,这与他人的研究结果相反,大多数的研究表明,DO不足会影响硝化作用的进行从而使氨氮去除率减低(Moss,1990;Fan et al.,2013b)。这可能是因为浮床植物的的存在可大大提高氮去除率,植物可以通过直接吸收作用、根系泌氧以及为微生物提供附着载体等途径提高净化效率(Ellis et al.,1994;Jenssen et al.,1993)。由于微生物活性与温度密切相关,冬季微生物活动减弱,故植物对于冬季人工湿地的净化效果尤为重要(Zou et al.,2016;Fan et al.,2016)。Tanner(1996)研究证明相对于无植物的人工湿地系统,植物生物量大的系统对氮的去除率更高,且植物对氮的吸收作用与植物生物量呈线性关系。本研究中,有较高生物量的 FCW-L氨氮和总氮浓度远低于 CK,FCW-L和CK植物吸收对氮的去除率分别占总氮去除率的12.14%和1.63%。FCW-L的水芹菜在后两个周期的生物量远大于前两个周期,相应地后两个周期的氮去除率增加,这与Wu et al.(2011)的研究结果相一致。
微生物途径的硝化与反硝化作用是氮去除的主要机制(Gottschall et al.,2007)。浮床植物根系泌氧能够改变根系微环境中的氧气状况,形成微氧环境,为好氧微生物提供适宜的生境,从而有利于硝化作用的进行(Wang et al.,2016)。Bodelier et al.(1996)研究表明,缺氧环境中植物根系泌氧可以为氨氧化细菌提供好氧生态位。浮床人工湿地与传统人工湿地相比缺少基质,植物根系成为微生物附着的主要载体(Zou et al.,2016;Wu et al.,2011)。植物根系是活跃的吸收器官和合成器官,根的生长情况和代谢水平直接影响植物地上部的生长和营养状况(黄磊等,2014)。本研究中,CK的植物生长缓慢,后期出现枯萎,FCW-L的水芹菜虽然单位根系活力较低,但叶绿素含量升高,尤其是后两个周期温度升高,水芹菜生长迅速,生物量和株高尤其是根系部分大大增加,保证了水芹菜的生长活性。同时,大量的水芹菜根系不仅可以改善系统的氧环境,还可为微生物提供丰富的生存空间,浮床植物根系表面细菌数量密集,氮去除率增高(Brix,1994;Tanner et al.,2003)。因此,添加植物秸秆不会对人工湿地造成二次污染,相反,添加植物秸秆配合浮床植物种植可明显提高冬季氮去除效果。
4 结论
本研究表明,添加芦苇秸秆:(1)可以促进冬季浮床人工湿地中水芹菜的生长,与 CK相比,FCW-L的水芹菜叶绿素增加3.8倍,总根系活力和生物量显著提高;(2)缓解尾水湿地中ω(C)/ω(N)比失衡问题,FCW-L的ω(C)/ω(N)平均值可达到13.1;(3)水芹菜浮床人工湿地配合添加芦苇秸秆可以提升冬季氮净化效果,与CK相比,FCW-L的NO3--N、NH4+-N和TN去除率分别提升了63.5%、41%、46.241%。研究结果对于实际工程应用中提高尾水湿地冬季净化功能及湿地植物的管理具有理论指导意义。
猜你喜欢
水产养殖(2022年2期)2022-11-05
土壤学报(2021年4期)2021-09-10
科学大众(2021年9期)2021-07-16
绿色科技(2020年24期)2021-01-09
生态学报(2019年8期)2019-05-31
基层中医药(2018年7期)2018-12-06
娃娃乐园·综合智能(2018年11期)2018-07-10
福建畜牧兽医(2016年5期)2016-12-16
环境科技(2016年1期)2016-11-08
西藏大学学报(自然科学版)(2015年1期)2015-12-25
