
基因沉默机制的基因表达式编程
2018-12-04 02:14:06张国锋刘丽萍
计算机工程与应用 2018年23期
郭 勇,张国锋,刘丽萍
1.黔南民族师范学院 计算机与信息学院,贵州 都匀 558000
2.黔南民族师范学院 生命科学与农学院,贵州 都匀 558000
1 引言
受生物界进化、学习、觅食等现象和行为的启发,衍生了众多的智能搜索算法,如遗传算法(Genetic Algorithm,GA)[1]、蚁群算法(Ant Colony Optimization,ACO)[2]、人工鱼群(Artificial Fish-Swarm Algorithm,AFSA)[3]、狼群算法(Wolf Pack Algorithm,WPA)[4]等,基因表达式编程(Gene Expression Programming,GEP)[5]是继GA之后受基因遗传理论启发的另一种智能搜索算法。公开GEP文献中大量使用形如的多项式函数进行实验佐证,实验表明多基因染色体GEP对多项式函数的挖掘具有较好的计算效果。在使用文献[6]中包含了正弦函数sin x、自然对数函数ln x、平方根函数Sqrt(x)等1目运算符的非多项式函数作为测试函数时,传统多基因染色体GEP的计算效果并不理想。为提升GEP对非多项式函数的挖掘能力,借鉴转基因工程中的基因沉默现象,定义新的多基因染色体结构,增加表达因子域,遵循“深度优先”原则对染色体中的基因进行连接解码,利用表达因子运算符的不同目数实现对染色体基因表达的抑制机制,通过遗传算子中基因转座算子和新的表达因子域突变算子,产生基因表达的位置效应,拓展出一种基因表达式编程算法——SFGEP(Gene Expression Programming of Symbol Field,SFGEP),SFGEP对包多种运算目数的非多项式函数的挖掘能力得到显著提升。
2 GEP与基因沉默简介
2.1 GEP简介
GEP以种群(Population)来模拟生命物种的遗传进化,种群由若干个体(individual)组成,个体抽象为染色体(Chromosome),染色体由1个或多个基因(gene)组成,基因编码字符集分操作符集F和终结符集T,基因的线性编码称为基因型,分为头部(head)和尾部(tail),头部编码取自F⋃T,尾部编码取自T,头部长度h和尾部长度t需满足关系式(1)。

式(1)中的M为运算操作符集F中的最大操作符目数。
基因编码按照“广度优先”原则构建的表达式树,称为表现型,对表现型表达式树后序遍历所得数学表达式称为基因解释。如F={+,-,*,/}和T={a,b},头部长度为6的基因型编码“−+b*/aabbbabb”,其表现型表达式树如图1。
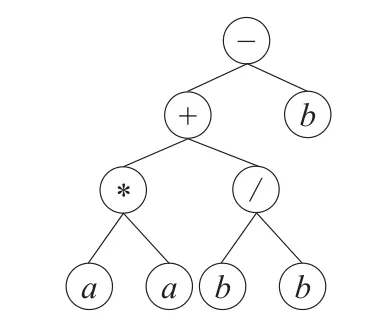
图1 表达式树
GEP通过选择、突变、转座、重组等遗传算子实现种群的进化,通过适应度及选择算子来控制种群演化方向。适应度函数可根据具体GEP问题进行设计,符号回归问题常用绝对误差、相对误差等统计函数作为适应度函数[7]:
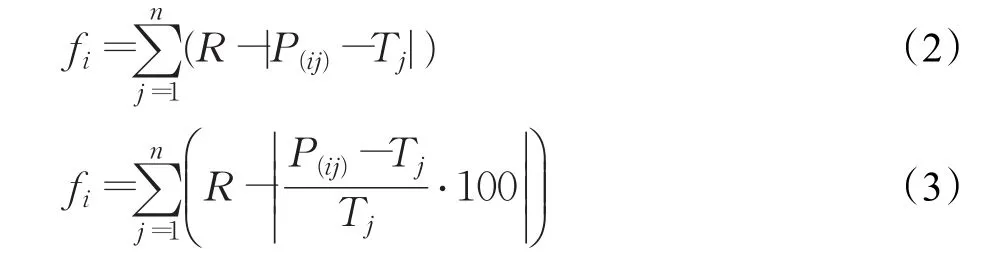
2.2 基因沉默
基因沉默(gene silencing)[8]指生物中特定基因由于种种原因不表达或者表达减少的现象,是基因表达调节的一种重要手段。基因沉默现象最早在植物转基因工程中受到关注,也称转基因失活。转基因沉默分为转录水平与转录后水平两种[9],其中转录水平的转基因失活有两种情况,一种情况是导入的外源基因植入宿主基因组时,由于位置效应,外源DNA位于转录的不活跃区,产生转基因的异染色质化,从而出现不转录或者低水平转录。另一种情况是转基因反向重复区形成茎环结构,重复同源拷贝数增加,出现甲基转移酶异识别、甲基化,导致基因转录无法进行。
基因沉默的现象不仅存在转基因工程中,在生物界也存在类似现象。如植物抗病毒侵染的自然机制——病毒诱导的基因沉默(Virus-Induced Gene Silencing,VIGS),又如返祖遗传中祖先部分原有性状的基因仍然存在,长期的进化使其不具备活性而沉默,由于多基因调控等因素而激活。转基因沉默可以应用于植物抗病的基因工程和植物功能基因组研究。基因沉默和激活沉默基因的技术具有正反两方面的现实意义,一方面在保留优良性状的生物基因工程中需要克服基因沉默现象,使优良基因得以表达,或者对生物原有优势性状的沉默基因进行激活。另一方面可利用基因沉默屏蔽原有的病理基因或劣势基因的表达。
3 基因沉默机制的SFGEP
3.1 GEP的“随机性”和“冗余性”
“随机性”是自然界演化普遍存在的现象,GEP作为一种仿生智能搜索算法,GEP遗传算子的随机性,使较小规模的种群可演化出更多可能模式,赋予了种群搜索GEP问题解的能力,保持一定的“随机性”是智能搜索算法有效的基础。在GEP基因中存在类似位置效应而无法在GEP表现型中得以表达的编码区域——中性区域,视为GEP基因编码中的“冗余”现象,这种“冗余”因遗传算子的实施而带来更多演化结果的可能性。
传统多基因染色体GEP中,采用固定连接运算符进行多基因的连接解释,染色体结构中基因个数固定,且所有基因在染色体的解释中都得以表现激活,不具备“冗余”情况,也不具备某个基因不表达的“沉默”现象,演化的“随机性”较弱。
3.2 SFGEP的染色体编码结构
SFGEP基因编码字符集包含操作符集F和终结符集T。SFGEP染色体结构由表达因子域S和表达基因域G两部分组成,如图2所示,表达因子域S的编码全部或部分取自操作符集F,S的符号集记为F′。
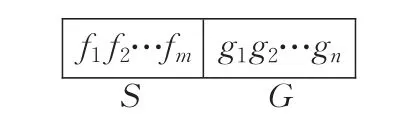
图2 SFGEP染色体结构
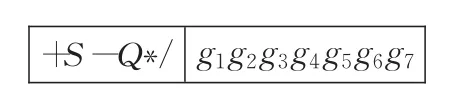
图3 SFGEP的染色体结构
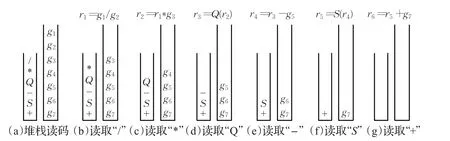
图4 SFGEP染色体的堆栈解释
图2 中 fi表示1个运算操作符,称为表达因子,gi表示单个基因,gi的编码结构与传统GEP相同。S中表达因子个数m与G中基因个数n的数量关系满足式(4):

式(4)中的 M 为F的最大操作符目数。式(4)保证SFGEP解码有足够基因满足S中所有表达因子运算的需要,使SFGEP染色体的结构具备完备性。
3.3 SFGEP的遗传算子
SFGEP的遗传算子包括针对表达基因域与传统GEP相同的遗传算子:突变(单点、两点)、转座算子(IS、RIS)、基因转座算子、重组算子(单点、两点)。SFGEP增加了1种遗传算子:表达因子域S的突变算子,简称为S突变,即在表达因子域S中随机选择1个突变位,随机取F′中1个符号进行替换。
3.4 SFGEP染色体解释
(1)单个基因的解释
SFGEP表达基因域G中单个基因的解释与标准GEP相同,遵循“广度优先建树”、“后序遍历表达式树”等原则进行解释,基于“建树解释”算法时空复杂度高,可使用文献[10]等“无树解释”算法,提升基因解释效率。
(2)染色体基因的连接解释
SFGEP对G中基因采用文献[11]“深度优先”的堆栈解释方式,由于S中的表达因子运算目数可能不相同,基本原则是“每个表达因子 fi都要从基因栈中弹出至少1个基因”,当 fi(i>1)运算目数为1时,不需要弹出的基因参与解释运算,则丢弃此基因,从而使此基因在解释中没有得到表达,成为“沉默基因”。
例1如图3所示的SFGEP染色体,表达因子域S部分的编码为6个运算操作符:+S-Q*/,表达基因域G包含7个基因,即关系式(4)取等号的情形。
例1中的SFGEP染色体基因连接堆栈解释过程如图4。图4(d)和(f)中由于表达因子Q和S的目数为1,不需要弹出的基因g4和g6参与运算,即g4和g6没有参与染色体的解释,形成基因沉默现象,显然沉默基因位受到了表达因子域中表达因子目数和位置的影响。
图4(g)中表达因子栈为空是染色体基因连接解释的终止条件。由于本例中表达基因域G基因个数取3.2节关系式(4)的“=”情况,所以表达基因栈同时为空,若关系式(4)取“>”,则基因栈中会存在冗余基因,这些冗余的基因与传统GEP基因的中性区域编码一样,可因“基因转座”算子等原因在演化中得以表达,形成另一种位置效应。例1表达因子域S对表达基因域G中的基因“激活”解释所得表现型如图5。
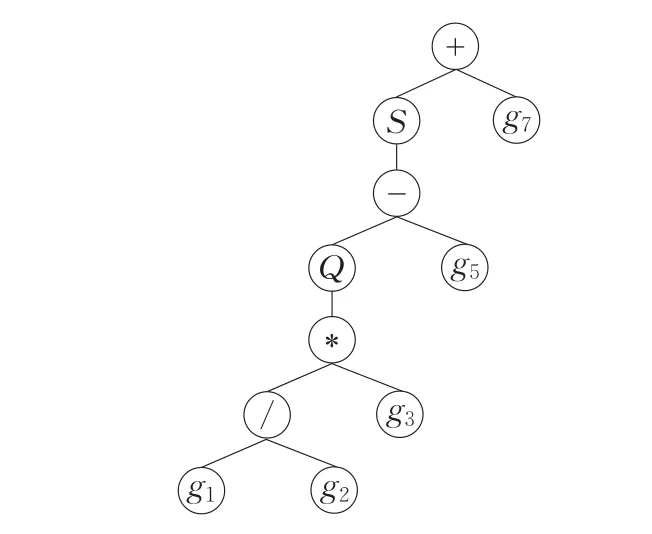
图5 SFGEP染色体基因的解释
4 对比实验与分析
本章以GEP公开文献及文献[6]中部分函数进行SFGEP与传统多基因染色体GEP函数挖掘的对比实验,实验一的表达因子符号集F′与操作符集F相等,统计标识为SFGEP-I,实验二限制表达因子符号集F′取F中部分符号,统计标识为SFGEP-II。
4.1 实验一
目标函数10个,其中前5个为公开文献中常用的多项式函数,后5个为包含了1目和2目运算符的函数,在对应取值范围内随机生成20组精度为10-16的训练样本。
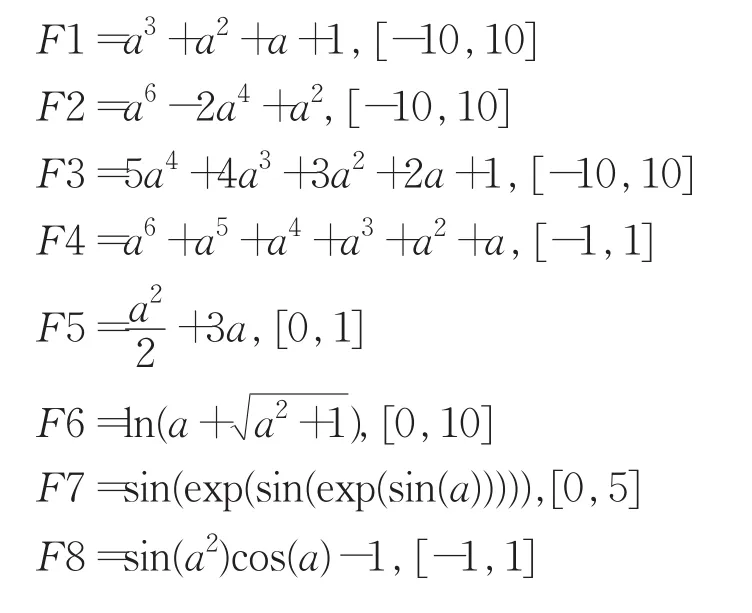

表达因子域长度取3,关系式(4)取“=”,所以表达基因域由4个基因构成。传统多基因GEP染色体也由4个基因构成,采用“+”作为染色体基因连接运算符,函数F3的基因头部长度为15,其他函数的基因头部长度均为10,表达因子符号集F′=F。适应度函数采用反映训练样本集整体拟合度的统计函数R2:

其中m为样本数,yi为样本集第i个样本函数,y′i为染色体对i样本的解析值,yˉ为所有样本解析的平均值。
实验适应度控制阈值取0.999 9,S突变率为0.1,其他遗传算子参数设置如表1。
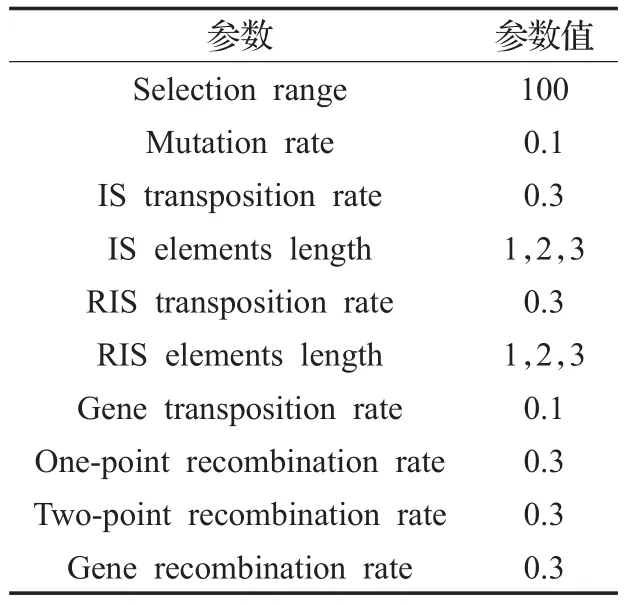
表1 遗传算子参数设置
演化种群大小为100个染色体,控制演化最大辈数为5 000辈,使用C#设计程序对每个目标函数分别使用SFGEP和GEP连续演化100次。实验统计项如表2。
(1)对于前5个多项式函数,传统GEP整体计算效果优于SFGEP,两者的成功率无显著差异。其原因主要在于本实验中GEP采用“+”作为多基因染色体的连接运算符,连接运算结果:g1+g2+g3+g4,形式上与多项式函数相近,GEP演化成功辈数的3项指标整体优于SFGEP。但由于“+”满足交换律,尽管传统GEP多基因的连接运算采用“广度优先”原则如图6(a),其运算结果与采用“深度优先”原则如图6(b)的结果相同,实验中的操作符集F均含有“+”,显然SFGEP的表达空间包含了GEP的表达空间,SFGEP随机演化搜索到最优解的概率相对较小,表现在平均成功辈数上SFGEP劣于传统GEP。
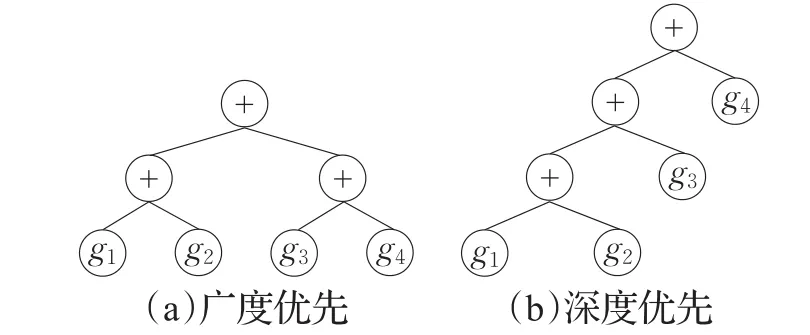
图6 多基因染色体连接解释
(2)对于后面5个包含有1目和2目操作符的非多项式函数,除了相对简单的函数F10外,其他略复杂的非多项式函数,SFGEP的整体计算效果明显优于传统GEP,主要体现在成功率方面。特别是在F7、F8、F9这三个函数的成功率上,SFGEP大幅领先于传统GEP。对于类似F10这样形式较简单的函数,SFGEP则不具备优势。
4.2 实验二
针对实验一中F10等函数的平均成功辈数较弱于传统GEP的结果,采取表达因子符号集F′取操作符集F的部分符号的措施再进行对比实验,即限制表达因子符号集F′的方法,并增加了以下五个目标函数:
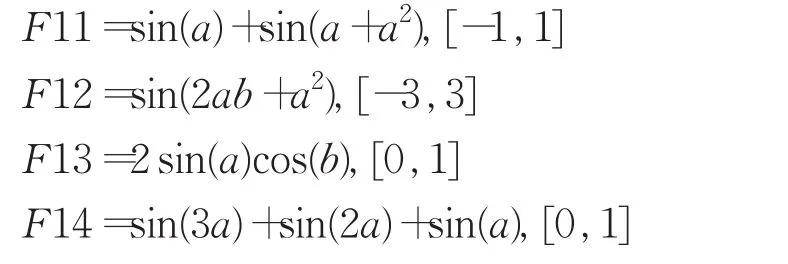
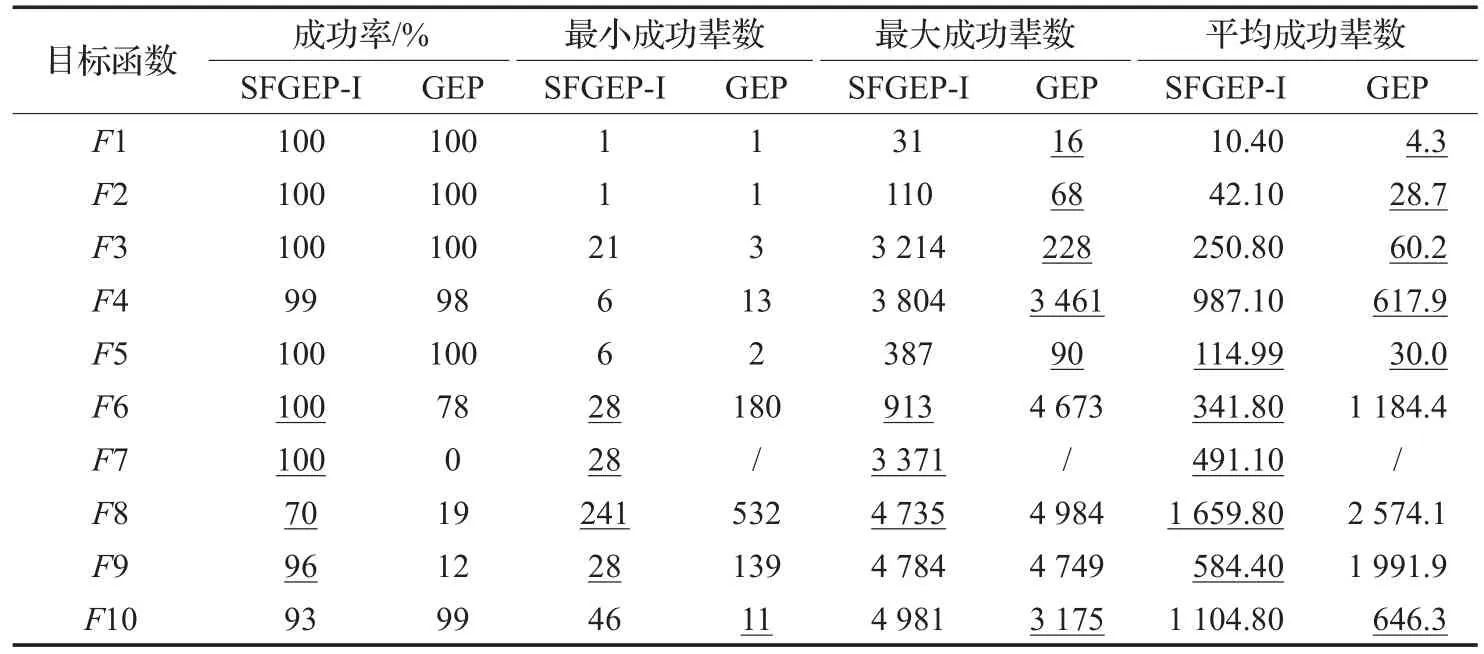
表2 实验一数据统计
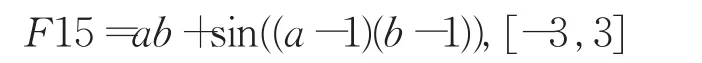
限制表达因子符号集F′的基本原则:(1)基于“+”与“-”、“*”与“/”的逆运算关系,二者取其一;(2)基于基本三角函数间的可换算关系,正弦函数、余弦函数、正切函数、余切函数四者取其一;(3)其他函数适当取舍,保持表达因子符号集F′至少包含2个符号。各目标函数的表达因子符号集F′如表3。

表3 实验二表达因子符号集F′设置
演化的其他设置与实验一相同,实验主要统计了SFGEP-II与传统GEP的成功率和平均成功辈数(四舍五入保留2位小数),成功率统计结果如表4。
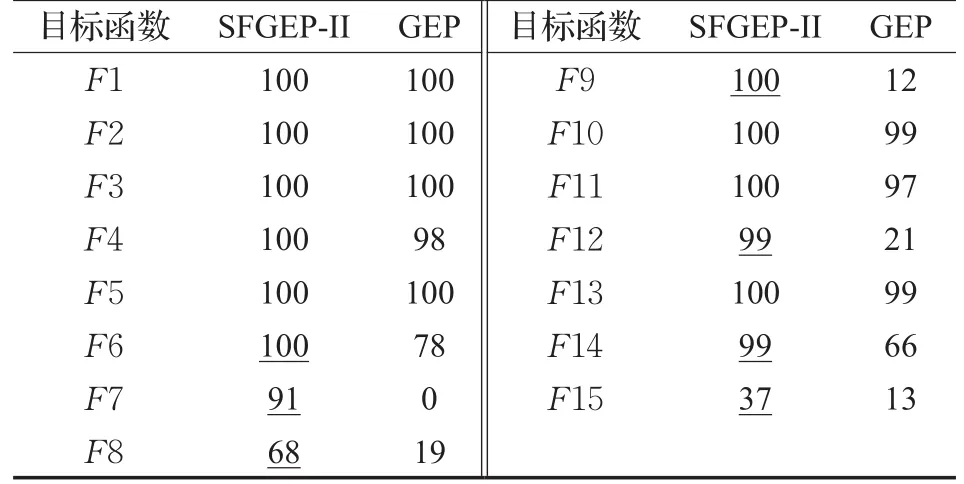
表4 实验二成功率统计%
表4数据表明,前5个多项式函数,SFGEP-II与GEP的成功率无显著差异。后10个非多项式函数SFGEP-II成功率整体比GEP高,7个带下划线的数据具有非常明显优势。平均成功辈数统计结果如图7。
图7(a)为多项式函数 F1~F5对比。SFGEP-II在平均成功辈数上与传统多基因染色体GEP仍有一定差距,但相较SFGEP-I来说差距大幅缩小,F1和F2基本无差异,F4反而是SFGEP-II的平均成功辈数更小。
图7(b)为非多项式函数F6~F15的对比。由于GEP对F7的成功率为0,因而没有列出。统计结果显示,SFGEP-II在平均成功辈数上都优于传统多基因染色体GEP,体现了SFGEP对非多项式函数的总体收敛速度更快。
图7(c)为非多项式函数的SFGEP-I与SFGEP-II的对比。除F7外,其余函数的SFGEP-II SFGEP-I的平均成功辈数小,这主要得益于适当的表达因子限制使SFGEP-II的表达空间有所缩减。
5 SFGEP效能分析
5.1 SFGEP的计算复杂度分析
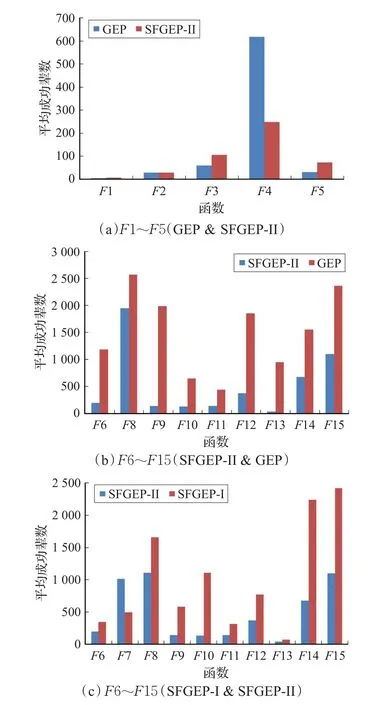
图7 实验二平均成功辈数统计
由于SFGEP染色体结构中包含了表达因子域S,在SFGEP染色体的基因个数取式(4)中的大于符号时,SFGEP的空间复杂度略高于传统多基因GEP,对巨大种群应用情景有一定影响,在没有突破硬件内存极限的应用环境中影响较小。
GEP的每代演化都要在样本集上对种群所有个体进行解码计算,其中染色体的每个基因的解码总耗时最多。对于符号回归问题,由于传统GEP一般采用运算目数为2的加号对染色体中多个基因进行连接解释,传统GEP多基因染色体中的所有基因都需要进行基因解码计算。SFGEP在进行染色体解释的过程中,可能因为表达因子S中存在运算目数为1的运算符,形成部分沉默基因,染色体解释不需要对这些沉默基因进行解码计算。尽管SFGEP演化中多了1个S突变算子,由于一般其突变率较小,每代演化仅进行1次S突变,而每代演化基因解码的计算量要大得多,加上多数符号回归问题只包含1目或2目运算符,染色体结构相近的SFGEP由于存在基因沉默现象,不需解码沉默基因,需要解码的基因总量相对传统GEP更少。所以,从整体演化计算过程来看,在种群规模相同,染色体结构基因数相近的情况下,SFGEP的计算时间复杂度不高于传统多基因染色体GEP。
传统多基因染色体GEP采用广度优先原则连接解释多基因染色体的方式,当不采用“+”(或“OR”)等具备交换律特性的连接符时,算法设计的时空复杂度较高,SFGEP采用深度优先原则连接解释多基因染色体,算法可使用时空复杂度更低的简单循环结构实现。
5.2 算法有效性分析
由于传统多基因染色体GEP采用固定连接符号“+”(或“OR”)进行基因连接运算,使得从染色体结构“模式”更接近于“多项式函数”,在多项式函数发现方面具有先天优势。SFGEP利用表达因子域中的多种符号进行基因的连接运算,染色体结构“模式”较接近于非多项式函数,虽在多项式函数发现上有一定弱势,但对第4章实验中的非多项式函数的发现有明显的优势。SFGEP既保持了对多项式函数的挖掘能力,同时具有良好的多种目数运算符、非多项式函数挖掘的效能。表达因子域的多操作符连接运算与“冗余”的基因域,使SFGEP可能包含更多形式的数学模型,从而带来更多的选择与可能,采用SFGEP有可能找到符合生产实际的数学模型,所以SFGEP具有一定的工程实践意义。
SFGEP对非多项式函数符号回归问题的有效性不仅因为“模式”上的优势,还来源于SFGEP采用深度优先原则连接多基因,相对于传统GEP的广度连接,SFGEP染色体的表现型表达式树更高,按照文献[12]基于表现型的GEP解空间模型理论,在更高的表现型子空间中等价的最优解接近于几何级数的增长趋势,相同基因数的染色体,SFGEP比传统GEP探索更高子空间的能力更强,根据文献[13-14]理论,随着演化代数的增加,种群中适应度更高的个体更多,SFGEP搜索到等价最优解的概率更大。
6 结束语
GEP给出了一种非二进制编码遗传进化算法的基本思想与计算框架,在此基础上许多GEP研究者针对解决不同类型问题和GEP效能的改进进行了GEP算法的拓展[15-20],SFGEP给出了一种包含表达因子域的多基因染色体结构和“深度优先”、“多表达连接运算符”解释染色体的GEP拓展计算框架,为解决生产实践中的GEP问题提供了一种新计算方案的选择。今后将进一步对SFGEP的生产实践应用、染色体基因数对演化效果的影响及调控、表达空间模式及其多解特性等方面深入研究。
猜你喜欢
中学生数理化(高中版.高考数学)(2022年1期)2022-04-26 14:09:30
数学物理学报(2021年2期)2021-06-09 08:54:26
应用数学(2020年2期)2020-06-24 06:02:44
数学小灵通(1-2年级)(2020年6期)2020-06-24 05:57:54
科学之谜(2019年3期)2019-03-28 10:29:44
数学年刊A辑(中文版)(2018年2期)2019-01-08 01:59:54
科学之谜(2018年8期)2018-09-29 11:06:46
中学生数理化·八年级数学人教版(2017年2期)2017-03-25 16:12:51
中学生数理化·七年级数学人教版(2016年9期)2016-12-07 08:18:09
数学物理学报(2016年3期)2016-12-01 05:36:27
