
向海地区斑嘴鸭体内1新种绦虫的分离与鉴定1)
2018-11-30 05:29:06侯志军王丽新沈美英柴洪亮杨思远
东北林业大学学报 2018年11期
侯志军 王丽新 沈美英 柴洪亮 杨思远
(东北林业大学,哈尔滨,150040) (黑龙江民族职业学院)
斑嘴鸭(Anaspoecilorhyncha)在向海地区很普遍且分布很广[1],隶属鸟纲、雁形目、鸭科、鸭属,中型游禽[2]。在我国东北部、北部及华北等地区繁殖,在我国长江中下游、东南沿海和台湾越冬。由于过度猎取和环境恶化,斑嘴鸭数量日趋减少,因此被列入《世界自然保护联盟》。
绦虫隶属于扁形动物门的绦虫纲。绦虫种类众多,生物学特性各异,分布广泛,对人和动物危害严重。鸟类[3],特别是水禽,可以寄生多种绦虫[4],引起严重的鸟类疾病。寄生于鸟类肠道内的绦虫会争夺宿主营养,或钻入宿主肠黏膜破坏肠壁组织,或由多数虫体密集造成肠阻塞,阻碍宿主的消化吸收而影响鸟类的正常生长发育[5],如戴文科的节片戴文绦虫(Davaineaproglottina)、膜壳科的毛形剑带绦虫(Drepanidotacnialanceolata)、片形皱褶绦虫(Fimbriariafasciolaris)等[6]。
膜壳科(Hymenolepididae)绦虫在哺乳动物和鸟类中有着广泛的寄生,如鼠类、蝙蝠和水禽等。目前至少有620种绦虫和膜壳科的230种绦虫寄生在鸟和哺乳动物中。面对如此多种类的绦虫,对它们分类问题的研究从未停止过。近期,Khalil et al.对膜壳科进行了比较系统的修订[7],但并没有对每一个属进行修正。膜壳科的绦虫种类繁多,部分种类形态学特征相似,如果仅仅依据形态学特征进行种属鉴定,由于缺乏明确形态学特征往往会导致物种分类鉴定的错误[8]。例如,在鸭小肠内分离出的1种膜壳科的绦虫(MonosaccanthesCzaplinski,1967),直到2001年才被发现是EchinatriumSpassky,1956的同物异种[9]。
核苷酸序列数据成功应用于寄生虫分类,这有助于系统解决膜壳科寄生虫分类面临的问题[10]。基于一个完整的形态特征评估的和系统发育的证据[11],曾被列入啮壳属(Glires)一些物种已被转移到最近成立的Pararodentolepis属[12]。此外,Greiman et al.在结合了核酸序列(28S,nad1)和形态学数据的基础上创立了一个新属[13]。显然,膜壳科分类进一步的完善需要形态学和分子数据的结合使用[14]。
在中国雁形目绦虫的研究非常少,目前为止还未见斑嘴鸭体内膜壳科绦虫的研究。
1 材料与方法
样品材料:斑嘴鸭取自吉林省向海地区。绦虫取自斑嘴鸭小肠内,置于70%酒精内用于形态学和分子研究。
镜检方法:将经苏木素染色[15]的绦虫标本在光学显微镜下进行形态学观察。
DNA的提取:取5 g液氮中保存的虫体组织,置于研钵中,研磨粉碎,使用组织细胞基因组DNA快速提取试剂盒提取DNA,-20 ℃下保存。
PCR扩增3个基因所用引物:扩增28S rDNA使用的引物参考Hillis et al.[16]设计的,目的片段长度为1 100 bp。扩增nad1基因使用的引物参考Widmer et al.[17]设计的,目的片段长度为700 bp。扩增18S rDNA基因引物设计是通过NCBI数据库检索相关18S基因序列,自主设计,目的片段长度为500 bp。所有引物序列均委托库美生物有限公司合成。引物序列详细见表1。

表1 28S rDNA、nad1、18S rDNA基因扩增引物序列
PCR扩增3个基因所用反应体系及反应条件:PCR扩增采用25 μL PCR体系,PCR buffer 2.5 μL,MgCl22.0 μL,dNTP 2.5 μL,Primer1 1.0 μL,Primer2 1.0 μL,DNA 3.0 μL,蒸馏水12.0 μL,Taq酶1.0 μL。28S序列PCR反应程序为,94 ℃预变性5 min;94 ℃变性30 s,58 ℃退火30 s,72 ℃延伸1 min,30个循环;72 ℃延伸7 min。nad1序列PCR反应程序为,94 ℃预变性5 min;94 ℃变性30 s,45 ℃退火30 s,72 ℃延伸50 min,30个循环;72 ℃延伸7 min。18S序列PCR反应程序为,94 ℃预变性5 min;94 ℃变性30 s,50 ℃退火30 s,72 ℃延伸30 s,30个循环;72 ℃延伸7 min。最后,将PCR产物在1%琼脂糖凝胶电泳下进行观察。
PCR产物测序及分析:用杭州博日科技有限公司的胶回收试剂盒纯化扩增产物,将纯化后产物连接到pMD18-T载体上,再将连接产物转化至感受态细胞中,振荡培养后涂布在含Amp的LB上,过夜培养后挑选白色的单菌落,对菌落进行PCR鉴定,筛选阳性克隆并经凝胶纯化后送至库美生物有限公司进行双向测序。用序列分析软件对测序结果进行排列校对,然后与GenBank数据库中的已知序列进行Blast比对,确定虫种。
系统进化分析:用软件Mega 5(http://www.megasoftware.net/)中Neighbor-Joining(NJ)法构建基于3个基因位点(28S rRNA、nad1和18S rRNA)序列的系统树,并用Kimura two-parameter模型计算遗传距离;设置1 000次bootstrap检测支持度。
2 结果与分析
2.1 形态描述
中型绦虫,虫体全长93.0 mm,体最大宽度1.4 mm。前端细小,后端渐大,节片数目多,具缘膜,所有节片均长远大于宽。头节圆形,两侧略膨大,宽度为0.391~0.412 mm,平均为0.402 mm(图1A)。吸盘4个,具棘刺,呈圆形或椭圆形,大小为(0.098~0.103)mm×(0.101~0.144)mm,平均为0.101 mm×0.122 mm。顶端有顶突,平均大小为0.171 mm×0.131 mm,其周围有一圈小钩,共10个,呈“y”字形,吻钩长0.019~0.029 mm(图1B)。
颈节与头节清楚地分开,节片宽为头节的一半,宽度为0.236~0.337 mm,平均为0.286 mm。
未成熟节片细小,其内生殖器官尚未发育成熟。
成熟节片大小为(0.901~1.208)mm×(0.079~0.127)mm,平均为1.055 mm×0.103 mm。内含成熟生殖器官一套(图1C,D)。生殖器官为单套。生殖孔开口于虫体右侧,在每个体节侧缘前端1/3处。
睾丸2枚,成熟的睾丸呈椭圆形,直线排列于节片中央,紧凑并位于中间,与卵巢常有部分重叠。大小为(0.056~0.081)mm×(0.078~0.126)mm,平均为0.069 mm×0.102 mm(图1C)。具有外贮精囊,囊状呈椭圆形,一般位于排泄管附近,与阴茎囊相连接。平均大小0.102 mm×0.045 mm(图1C,D)。内贮精囊呈长梭形,占据阴茎囊的大部分,与阴茎囊几乎等长(图1C,D)。在成熟节片内,阴茎囊伸长但未到达节片中央,也并未穿过背渗透调节管,大小为(0.175~0.190)mm×(0.048~0.054)mm,平均为0.183 mm×0.051 mm(图1C,D)。
卵巢囊状位于中间,呈三叶,宽度为0.290~0.396 mm,平均为0.343 mm(图1D)。卵黄腺圆形或椭圆形,紧凑并位于中间,背对卵巢,大小为(0.071~0.096)mm×(0.056~0.095)mm,平均为0.084 mm×0.076 mm(图1D)。
孕卵节片(1.249~1.374)mm×(0.121~0.136)mm,平均为1.312 mm×0.128 mm(图1E)。背排泄管比腹排泄管细,腹排泄管直径0.029~0.045 mm,平均为0.037 mm。背渗透压管直径0.027~0.033 mm,平均0.029 mm(图1C,E)。孕节子宫呈囊状,囊袋不裂解为卵袋。子宫壁向节片四周扩张,越出排泄管,充满整个孕节(图1E)。
综上,分离的绦虫为1新种,命名为Diorchispoecilorhynchamorbus, Zhijun Hou,2017。
词源,拉丁语,morbus=病,poecilorhyncha=宿主的拉丁名;指的是该种绦虫对斑嘴鸭致病。模式样本存放于东北林业大学寄生虫研究中心,NEFU-HY1901,头节及部分节片染色及固定在加拿大树脂中(1条),NEFU-HY190P两条没有头节的样本染色及固定在加拿大树脂中(2条)。

A.头节;B.吻钩;C.雄性器官;D.雌性器官;E.孕节。
2.2 分子进化分析
2.2.1 28S rDNA进化树
Bayesian法构建的28S序列的进化树结果见图2。在Bayesian树中,所测绦虫基因序列与Gen-Bank数据库上的其他绦虫的28S序列建立系统进化树分析,假叶目绦虫(Paraechhinophallusjaponicas)作为外群,位于进化树最底部。其余样本另开一支,与外群距离远,表明其余样本不属于假叶目,而是圆叶目。在圆叶目大枝中,又大致可以将所检测样本归为较大的5个小枝,包括裸头科(Anoplocephalidae)、膜壳科(Hymenolepididae)、双壳科(Dilepididae)、戴文科(Davaineidae)和中绦科(Mesocestoididae)。本试验的绦虫归于膜壳科分枝中。

图2 Bayesian法构建的绦虫28S的系统进化树
2.2.2 线粒体nad1进化树
Bayesian法构建的nad1序列的进化树结果见图3。在Bayesian树中,所测绦虫基因序列与GenBank数据库上膜壳科中其他属绦虫的nad1序列建立系统进化树。遗传进化树分析结果表明,本试验的绦虫并未归于膜壳科中已有nad1序列的各属绦虫之中,而是同膜壳属、假裸头属、微棘属并列,单独分为1枝。从亲缘关系上看,与微棘属、假裸头属、膜壳属的遗传距离比较近。
2.2.318S rDNA基因的绦虫序列比对和遗传距离
根据18S rDNA测序返回结果,通过NCBI上数据库Blast程序进行同源对比,从GenBank查找并下载相近的9属11种膜壳科绦虫的18S基因序列(包括双睾属、绉缘属、膜壳属、微棘属、啮壳属、Wardoides属、Coronacanthus属、Diploposthe属、Staphylocystis属),连同本试验获得的12个序列的18S rDNA基因遗传距离的关系见表2。由表2可见,Diorchispoecilorhynchamorbus与双睾属的D.brevis遗传距离最近,为0.01。
3 结论与讨论
结合绦虫典型的形态学特征,如节片有缘膜且宽大于长、睾丸数量少(2个),生殖孔为单侧开口(右侧),28S rDNA进化树可以确定本研究的绦虫为膜壳科绦虫。
在28 rDNA进化树中,D.poecilorhynchamorbus与Hymenolepissp.的亲源关系最近;在nad1进化树中,其同微棘属、假裸头属、膜壳属的遗传距离较近,但是微棘属和膜壳属绦虫的吸盘无刺且睾丸为3枚,假裸头属与膜壳属绦虫头节没有顶突或顶突退化,而本试验绦虫有2枚睾丸且顶突明显。18S rDNA分析表明其同双睾属的Diorchisbrevis亲缘关系(相似度大于99%)最近。
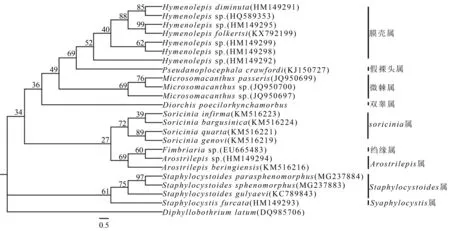
图3 Bayesian法构建的nad1的系统进化树

绦 虫1234567891011121 Diorchis brevis2 Fimbriaria sp.0.073 Hymenolepis nana1.071.064 Wardoides nyrocae1.101.070.035 Coronacanthus omissus1.071.060.010.036 Diploposthe laevis0.080.061.051.051.057 Microsomacanthus pachycephala1.061.040.020.040.021.038 Rodentolepis microstoma1.671.631.331.381.361.581.339 Staphylocystis furcata0.070.031.071.101.080.051.061.5710 Hymenolepididae0.070.041.051.061.050.051.031.610.0311 Hymenolepididae1.081.050.010.030.011.040.021.361.081.0412 D. poecilorhynchamorbus0.010.071.071.091.070.081.051.680.060.071.08
在膜壳科绦虫的形态学鉴定中,顶突及其棘刺、睾丸的数量等形态学特征是重要的依据[18]。本试验绦虫的头节上有具10个棘刺的可伸缩顶突以及每个节片具睾丸2个,符合双睾属的典型特征。综合形态学和分子进化特征,可确定该绦虫为膜壳科双睾属绦虫。
通过观察比较本试验的虫体,其基本特征与主要构造如阴茎囊的位置、阴茎囊的长度范围、睾丸位置形状等,同D.excentricusMayhew,1925相符。但是D.excentricus绦虫的棘突上的小钩长度为0.027~0.039 mm,而笔者描述的虫体上的小钩明显较小(0.019~0.029 mm),所以确定本研究分离的绦虫为膜壳科双睾属1新种,命名为D.poecilorhynchamorbus,Zhijun Hou,2017。
膜壳科是绦虫中种的数量最多的科[19],但该科绦虫的许多描述是不完整的,此外,膜壳科的分子数据也很少。本研究获得的试验数据,将补充到相关的数据库,为将来膜壳科绦虫更为科学的系统分类提供支持。
猜你喜欢
南方农机(2023年2期)2023-01-09 08:20:50
天津市教科院学报(2021年5期)2021-11-10 07:32:40
生物学通报(2021年9期)2021-07-01 03:24:44
奇闻怪事(2020年11期)2020-12-21 03:57:52
石河子大学学报(自然科学版)(2020年2期)2020-04-30 08:40:58
兵器装备工程学报(2019年1期)2019-02-25 10:02:52
中兽医学杂志(2019年5期)2019-01-06 11:53:51
中国全科医学(2017年14期)2017-05-24 14:46:44
江苏农业科学(2016年8期)2017-02-15 19:54:11
中国人兽共患病学报(2015年3期)2015-06-24 14:31:57
