
结核分枝杆菌拟核结合蛋白Lsr2阻遏调控rv1057基因转录的研究
2018-11-30 03:09:08付加芳张佩佩宗工理王新圆刘萌曹广祥
生物技术通报 2018年11期
付加芳 张佩佩 宗工理 王新圆 刘萌 曹广祥
(1. 山东省医药生物技术研究中心,济南 250062;2. 山东大学 微生物技术国家重点实验室,济南250100)
结核分枝杆菌(Mycobacterium tuberculosis)是肺结核病的病原菌,携带人口数约占全世界总人口的1/4,结核分枝杆菌多重耐药性菌株的发病率已经接近10%[1],因此结核病的预防和治疗面临严峻挑战。研究表明,结核分枝杆菌的致病性与其分泌蛋白有关,分泌蛋白参与了病原菌与宿主之间的相互作用,在结核分枝杆菌侵入、潜伏和致病等方面扮演着重要的角色。Type VII分泌系统是在结核分枝杆菌中发现的分泌系统,包括ESX-1、ESX-2、ESX-3、ESX-4和ESX-5等,负责一些与致病性有关的蛋白质分泌[2]。例如,ESX-1系统负责分泌ESAT-6和CFP-10蛋白,ESAT-6是在感染早期分泌的主要抗原蛋白,能够抑制巨噬细胞的自噬功能,帮助结核分枝杆菌从溶酶体逃逸到细胞质,诱导巨噬细胞的凋亡[3-6];而ESX-5系统负责分泌PE和PPE大分子蛋白,PE和PPE与细菌免疫原性有关,参与了修饰巨噬细胞成熟化、诱导促炎症因子IL-1β表达和诱导巨噬细胞死亡等过程[7-8]。
结核分枝杆菌有一层由分枝菌酸组成的外膜,在细菌表面形成通透性屏障阻碍蛋白质的分泌,现在还不清楚其外膜运输机制,例如ESX-1的外膜运输通道[9]。Rv1057是结核分枝杆菌中唯一的7-折叠片β-螺旋蛋白,β-螺旋蛋白具有典型的筒状蛋白质结构特征,生物功能包括转运蛋白、结构蛋白、信号蛋白和细胞膜蛋白等[10]。结核分枝杆菌在巨噬细胞内的生长早期大量合成Rv1057,而生长后期几乎检测不到Rv1057[11]。此外,敲除rv1057基因会显著降低ESAT-6的分泌[12],说明Rv1057可能参与了结核分枝杆菌的早期感染过程。
rv1057基因存在复杂的转录调控,其上游基因间隔区域具有1 003个碱基。前期研究显示rv1057基因受到双组份信号转导系统(Two-component regulatory system,TCS)MprAB和 TrcRS的 调 控,应答调控蛋白TrcR可以结合rv1057启动子区域中富含AT碱基的序列[11],MprA在rv1057启动子区域有多个结合位点[13]。MprA是激活rv1057转录的调控因子,TrcR则是阻遏rv1057转录的调控因子[14]。
原核生物染色体形成直径仅数微米的拟核结构,由一系列拟核结合蛋白(Nucleoid-associated proteins,NAPs)维持,Lsr2是革兰氏阳性菌中第一个被发现的拟核结合蛋白[15],在低氧、营养缺乏等压力条件下,lsr2基因表达明显上调,并通过与DNA结合形成物理屏障,协助细菌对抗损伤[16]。研究发现Lsr2在结核分枝杆菌基因组上有数百个结合位点,说明Lsr2能发挥全局性调控作用[17]。我们预测Lsr2可能是rv1057基因转录的一个阻遏蛋白,本研究通过凝胶阻滞迁移试验(Electrophoretic mobility shift assay,EMSA)分析和转录水平分析解析了Lsr2对rv1057基因的调控方式,研究结果可为阐明Rv1057的生物学功能提供试验依据。
1 材料与方法
1.1 材料
1.1.1 菌株和质粒 结核分枝杆菌野生型菌株H37Rv为本实验室保存,△lsr2突变菌株为Voskuil MI馈赠[16]。大肠杆菌 DH5α和 BL21(DE3)购自碧云天生物技术公司。pEASY-Blunt载体购自全式金生物技术公司。携带无启动子lacZ报告基因的pSM128载体和表达载体pET15b为本实验室保存。
1.1.2 实验试剂 7H9、7H10、OADC、胰蛋白胨、酵母提取物购自美国Difco公司,[γ-32P]ATP 购自美国PerkinElmer公司,T4 Polynucleotide Kinase购自美国Promega公司,Silica/Ceramic Beads购自美国MP公司,M-MLV购自Thermo公司,SYBR Premix EX Taq购自大连宝生物。
1.2 方法
1.2.1 重组Lsr2蛋白和TrcR蛋白的表达与纯化 以结核分枝杆菌H37Rv基因组DNA为模板,用PCR扩增Lsr2蛋白的编码基因,连接到pEASY-Blunt组成pBlunt-Lsr2,并进行DNA测序验证。测序正确后用NdeI和XhoI进行酶切,切下的片段连接到pET15b质粒构建重组表达载体pET15b-Lsr2。然后将pET15b-Lsr2转入大肠杆菌BL21(DE3),经IPTG诱导和Ni-NTA 柱(Sangon)纯化获得重组Lsr2蛋白,纯化的重组Lsr2蛋白经SDS-PAGE分析和浓度检测后备用。TrcR蛋白用已报道的方法进行制备[14]。Lsr2编码基因的PCR扩增引物为:Lsr2-15b-F:CATATGGCGAAGAAAGTAACCGTCACCT和 Lsr2-15b-R:CTCAGATCAGGTCGCCGCGTGGTATGCGTC。
1.2.2 EMSA分析Lsr2蛋白与rv1057启动子片段的结合情况 EMSA试验按照已报道的方法进行[14]。首先采用 Rv1057-ForN/Rv1057-RevN 引物PCR扩增含Lsr2结合位点的rv1057基因的255 bp(-503 - -249)启动子片段,并用[γ-32P]ATP和T4 Polynucleotide Kinase进行末端标记,然后与适量Lsr2蛋白溶液温育,8 %非变性聚丙烯酰胺凝胶电泳分离,最后用X-胶片曝光。
采用同样的方法分析Lsr2蛋白、TrcR蛋白与带突变位点启动子序列的结合情况,带突变位点的81 bp启动子序列采用化学合成获得。rv1057基因启动子片段的PCR扩增引物为Rv1057-ForN:TGACCACTAACCAGTCTCATCG,Rv1057-RevN:GGTATGTGGCAAACCAGTGCTA。
1.2.3 启动子-lacZ报告基因菌株构建和lacZ基因转录水平分析 报告基因菌株构建按照已报道的方法进行[12]。采用重叠PCR扩增不同区域和带突变位点的rv1057启动子片段,连接到中间载体并测序验证,然后连接到pSM128载体上,构建启动子-lacZ融合载体。重组质粒和pSM128空质粒通过Gene PulserXcell(BioRad)电击转化分别转入H37Rv菌株,用50 μg/mL链霉素筛选转化子并用PCR进行验证。
将导入不同重组pSM128质粒的H37Rv菌株培养至对数生长期,然后接入新鲜7H9/OADC培养基中并加入0.05%十二烷基硫酸钠(Sodium dodecyl sulfate,SDS),分别在第24 h,48 h和120 h收集菌体,提取总RNA,反转录成cDNA,并用荧光实时定量PCR(qRT-PCR)分析基因转录水平,试验重复3次。用不加模板的样品作为阴性对照,并用看家基因16S rRNA作为内标校正因样品初始浓度不同而造成的差异。
lacZ融合载体构建的PCR扩增引物:Rv1057-fusFor1:TGATCACTAGTGCTTGGTTTGTTCGCCG,Rv1057-fusRev1:TGATCAGCGCCGCTCCTCCTCATCA。lacZ基因上游:CCTGAGGCCGATACTGTCGT,下 游:TTGGTGTAGATGGGCGCAT;16S rRNA基因上游:TCCCGG GCCTTGTACACA,下游:CCACT-GGCTTCGGGTGTTA。
1.2.4 △lsr2突变菌株中Rv1057蛋白表达分析 将H37Rv和△lsr2突变菌株培养到对数生长期,转接到新鲜7H9/OADC培养基中并加入0.05% SDS,并在37℃水平摇床上晃动培养,分别在第24 h,第48h和120 h离心收集菌体。提取总RNA,反转录成cDNA,并用qRT-PCR分析lsr2基因转录水平。用不加模板的样品作为阴性对照,并用看家基因16S rRNA作为内标校正因样品初始浓度不同而造成的差异。同时用FastPrep(MP)振荡破碎菌体,离心收集上清,制备菌体蛋白。菌体蛋白按照以前的方法进行Western blot检测[14],检测Rv1057蛋白表达量。
2 结果
2.1 Lsr2对rv1057启动子的结合分析
Lsr2是一个拟核结合蛋白,生物信息学分析发现rv1057启动子中有一个可能的Lsr2结合区域。
EMSA实验结果(图1-A)显示,重组Lsr2蛋白在体外条件下可以特异性地结合rv1057启动子片段,提高Lsr2蛋白用量时阻滞效应更加明显,而过量的非标记rv1057启动子片段能够将标记的rv1057启动子片段从Lsr2蛋白上竞争性解离下来,表明Lsr2可以结合rv1057的启动子。与此同时,TrcR也结合该启动子片段,而且同时加入Lsr2和TrcR蛋白产生2条阻滞条带(图1-B),说明Lsr2和TrcR都能够结合rv1057启动子。
我们进一步设计了可能含突变TrcR或Lsr2结合位点的启动子序列M1-M4(图1-E),结果(图1-C和1-D)显示Lsr2和TrcR结合野生型rv1057启动子片段和M1片段,仅Lsr2能结合而TrcR不结合M2片段,仅TrcR能结合而Lsr2不结合M4片段,Lsr2和TrcR都不能结合M3片段。
2.2 Lsr2和TrcR对rv1057启动子的调控分析
本研究构建了含有野生型和突变型rv1057启动子的pSM128重组载体,分析rv1057启动子上Lsr2结合位点对报告基因lacZ转录的影响。
qRT-PCR结果(图2)显示,包含完整rv1057启动子的P1 可以正常转录报告基因lacZ,但第120小时lacZ转录水受明显受到抑制,第24小时lacZ表达量为第120小时lacZ表达量的6.25倍,差异有统计学意义(t=27.84,P<0.05);缺失Lsr2结合位点的P1-M4启动子也能正常激活lacZ的转录,但第120小时lacZ转录水平显著高于野生型rv1057启动子,是野生型启动子的8.94倍,差异有统计学意义(t=13.54,P<0.05);缺失TrcR结合位点(P1-M2)时lacZ转录水平在第24小时明显升高,第120小时与野生型rv1057启动子无明显差异;而同时缺失Lsr2和TrcR结合位点(P1-M3)时,lacZ转录水在第24小时和第120小时都显著高于野生型rv1057启动子,差异有统计学意义(t值分别为8.61和20.23,P<0.05)。
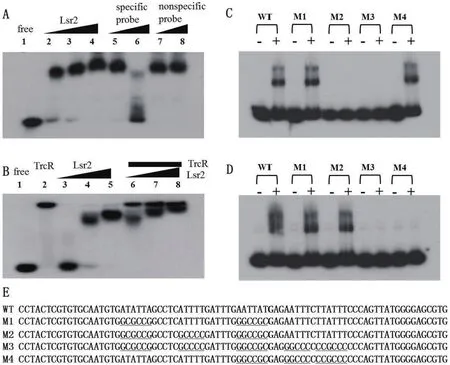
图1 Lsr2和TrcR结合rv1057启动子的EMSA分析
报告基因转录分析结果说明TrcR可能在感染早期发挥阻遏作用,而Lsr2可能在感染的中后期发挥阻遏作用。

图2 突变Lsr2位点对rv1057启动子的调控作用
2.3 Lsr2对rv1057表达的影响分析
为进一步研究在结核分枝杆菌内Lsr2对rv1057表达的影响,本研究采用qRT-PCR分析H37Rv和△lsr2突变菌株中trcR和lsr2转录水平的变化情况,并用Western blot分析Rv1057蛋白的表达情况。
qRT-PCR分析结果(图3-A)显示,培养24 h H37Rv菌株中lsr2转录水平较低,而第48小时和120 h的lsr2转录水平基本一致,分别是24 h的5.03倍和4.53倍,差异有统计学意义(t值分别为24.44、16.86,P<0.05);但是在H37Rv菌株中trcR转录水平的变化趋势与lsr2转录水平相反,第24小时的trcR转录水平是第48小时的4.86倍,是第120小时的10.03倍(图3-B),差异具有统计学意义(t值分别为6.79、10.16,P<0.05)。△lsr2突变菌株中trcR转录变化趋势与H37Rv菌株基本一致(图3-B)。以上实验结果说明TrcR主要在结核分枝杆菌生长早期表达,而Lsr2在结核分枝杆菌生长中后期表达。
Western blot分析结果(图4),显示,H37Rv菌株中Rv1057在培养24 h和48 h的表达量较高,培养120 h Rv1057表达量非常少,而△lsr2突变菌株在培养120 h的Rv1057表达量较高,说明Lsr2在结核分枝杆菌生长中后期抑制Rv1057的表达,与qRT-PCR试验的分析结果一致。上述结果表明Lsr2是rv1057基因的负调控因子,在结核分枝杆菌生长的中后期阻遏rv1057基因的表达。
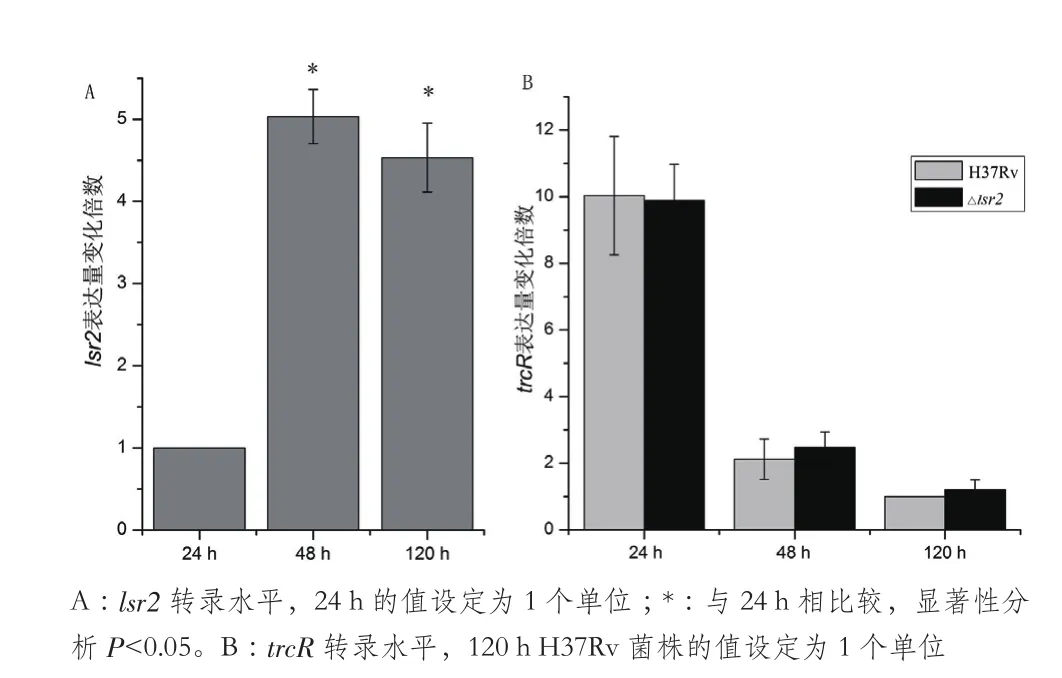
图3 lsr2和trcR转录水平的qRT-PCR分析
综合上述试验结果,本研究进一步完善了rvl057转录调控的模型:Lsr2和TrcR都是阻遏调控蛋白,MprA是激活调控蛋白;在结核分枝杆菌生长早期,TrcR结合rvl057启动子并阻碍rvl057的转录,当受到环境条件压力后MprA激活rvl057的转录并阻遏trcR表达,而随着时间的延长结核分枝杆菌开始表达Lsr2并阻碍rvl057的表达。
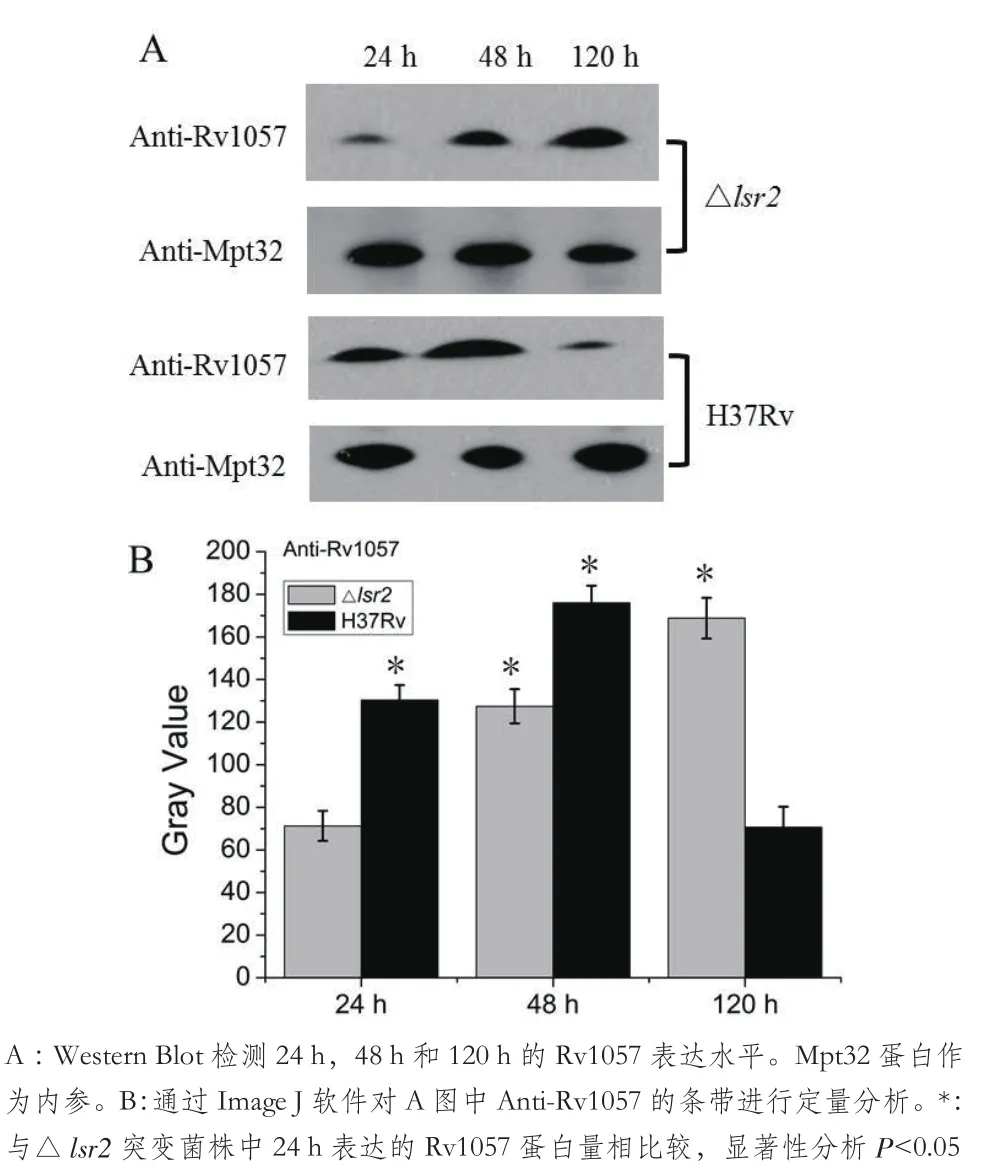
图4 H37Rv和△lsr2突变菌株中Rv1057蛋白的表达分析
3 讨论
Rv1057是一个7-折叠片β-螺旋蛋白,这类β-螺旋蛋白在细菌中具有功能多样性[10]。我们前期发现敲除rv1057基因影响结核分枝杆菌在巨噬细胞内的存活率[18],以及 ESAT-6和 CFP-10的分泌[12],说明rv1057可能影响结核分枝杆菌感染早期与巨噬细胞之间的应答,参与结核分枝杆菌与宿主之间的相互作用。转录组测序分析显示在抑制细胞合成药物万古霉素的压力下rv1057转录水平上升[19];Western blot检测显示在感染巨噬细胞的早期大量表达Rv1057,但感染后期检测不到Rv1057[11];我们课题组前期研究则表明Rv1057在结核分枝杆菌受到细胞膜压力诱导(例如SDS或Triton X-100)的情况下被大量表达[13],而且MprAB和TrcRS双组份信号转导系统同时参与rv1057的转录调控[13,18],这些研究结果提示rv1057受到了非常精细和复杂的调控作用。
Lsr2是一个拟核结合蛋白,在革兰氏阳性菌中广泛存在[15]。染色质免疫共沉淀和基因表达芯片实验表明,Lsr2能够结合基因组的高AT含量区域并调控基因转录[17]。研究显示,TrcR和Lsr2都结合富含AT碱基的DNA序列,但是TrcR和Lsr2的结合位点不同,Lsr2的 DNA结合结构域能够特异性结合在高AT含量区域的DNA双螺旋结构小沟,沿DNA小沟方向伸展并覆盖约5个AT碱基序列[20]。rv1057启动子存在高AT含量区域,研究显示TrcR结合rv1057启动子的高AT区域,并且阻遏rv1057的转录[11]。本研究证实Lsr2也结合rv1057启动子的高AT区域,与TrcR均可阻遏rv1057的转录,但两者在不同的生长时期发挥阻遏作用。本研究发现H37Rv菌株中Rv1057在培养48 h的表达量较24 h的表达量有所升高(图4),暗示rv1057除了受到Lsr2和TrcR的调控,还受到其它调控因子(比如MprAB双组份信号转导系统)的调控,说明rv1057的表达受到非常精细和复杂的调控作用。
综上所述,本研究发现Lsr2结合rv1057启动子的高AT含量区域,和TrcR均可阻遏rv1057基因的表达。但TrcR在诱导条件下可以完全阻遏rv1057基因的表达,而Lsr2可能起到基础阻遏作用,不能完全封闭rv1057基因的表达。
4 结论
本研究通过凝胶阻滞迁移实验、荧光定量PCR实验和Western blot实验发现结核分枝杆菌拟核结合蛋白 Lsr2是rv1057基因转录的阻遏蛋白,并且与已知的阻遏蛋白TrcR均可调控rv1057基因的表达;Lsr2在结核分枝杆菌生长周期的中后期大量表达并阻遏rv1057的转录,TrcR则在结核分枝杆菌生长周期的早期阻遏rv1057的转录。
猜你喜欢
华人时刊(2022年9期)2022-09-06 01:02:44
上海金属(2021年6期)2021-12-02 10:47:20
昆明医科大学学报(2021年3期)2021-07-22 07:40:04
华人时刊(2020年15期)2020-12-14 08:10:36
生物学通报(2019年3期)2019-02-17 18:03:58
特别健康(2018年4期)2018-07-03 00:38:26
中国医学装备(2016年6期)2016-12-01 06:44:41
广州大学学报(自然科学版)(2015年4期)2015-12-23 11:50:10
中国当代医药(2015年36期)2015-03-11 20:03:30
西南军医(2014年4期)2014-03-03 16:50:35
