
LAIR1基因3’端非翻译区多态性与胃癌发病风险的关系研究
2018-11-23 07:47林辉徐丹萍诸溢扬郑路亚
浙江医学 2018年21期
林辉 徐丹萍 诸溢扬 郑路亚
在我国,胃癌发病率居所有恶性肿瘤的前三位,且有明显的地域差异。胃癌发病年龄集中在50岁以上人群,且男女发病率明显失衡。人白细胞相关免疫球蛋白样受体 1(leukocyte-associated Ig-like receptor 1,LAIR1)几乎表达于所有免疫细胞,包括NK细胞、T细胞、B细胞、单核细胞、单核细胞衍生树突状细胞、嗜酸细胞、嗜碱细胞及肥大细胞等[1]。LAIR1的细胞质尾有2个ITIM基序,体外研究表明LAIR1通过单克隆抗体与分子间的交联反应,使细胞质尾的ITIM基序的酪氨酸磷酸化,再招募SHP-1、SHP-2和Csk发挥免疫抑制功能[2]。查询国外千人项目基因组测序数据库(IGSR)发现,中国西双版纳傣族与北京汉族人种LAIR1基因3’非翻译区(3’UTR)单核苷酸多态性(SNP)位点基因型分布频率存在明显差异。而本研究对胃癌与慢性胃炎患者LAIR1基因3’UTR的SNP位点基因分布频率进行分析,以探讨其与胃癌发病风险的关系。
1 对象和方法
1.1 对象 选取2014年10月至2015年9月在本院经胃镜下组织活检确诊的121例胃癌患者为胃癌组,其中男 91 例,女 30 例;年龄 30~87[64(57,71)]岁;肿瘤类型:鳞癌1例,腺癌95例,印戒细胞癌4例,腺癌+印戒细胞癌20例,神经内分泌癌1例。121例患者中92例接受胃癌根治术,明确肿瘤分化程度:中高分化41例,低分化51例;TNM分期:Ⅰ~Ⅱ期34例,Ⅲ~Ⅳ期58例;淋巴结转移阳性率为36.5%。选取同期本院收治且年龄、性别与胃癌患者匹配的126例慢性胃炎患者为对照组,均接受过胃镜检查,胃镜下可见椭圆形、圆形溃疡或糜烂,组织活检结果为慢性胃炎;其中男94例,女32例;年龄 35~75[60(51,65)]岁。在 IGSR 查询到傣族、北京汉族和南方汉族人种LAIR1基因3’UTR的SNP位点详细信息,见表1。本研究经医院医学伦理委员会审查通过。

表1 LAIR1基因3’UTR的6个SNP位点详细信息
1.2 一代测序法检测LAIR1基因3’UTR序列 取乙二胺四乙酸抗凝血,按照血液DNA抽提试剂盒操作说明书(GK1072,100T,上海捷瑞生物工程有限公司)对外周血全基因组DNA进行抽提。在保守区域设置引物,采用正向引物LAIR1F-5′-CCCACAGTCCACAAAGCCCAT-3′,反向引物 LAIR1R-5′-TGAGATTCTGCCGCCTCCTAGT-3′对 LAIR1 基因的 3’UTR 的 (19∶5355287-54355435)进行PCR扩增,PCR产物片段长度为1 212bp,经10%聚丙烯酰胺凝胶电泳进行确认。全部PCR产物经割胶回收纯化,同时在中间段设置测序引物 LAIR1M-5′-TGCGTGGAGTAAAGCACAACCC-3′,采用一代测序仪(Applied Biosystems)进行正向及中段测序。
1.3 序列比对 通过dbSNP数据库在19号染色体5355287-54355435位(LAIR1基因3’UTR)可查询到267个SNP位点,两组患者的测序结果采用序列比对工具BLAST进行比对,发现主要存在6个SNP位点,见表1和图1。
1.4 统计学处理 应用SPSS 17.0统计软件。胃癌组与对照组患者基因型及等位基因分布频率的比较采用χ2检验或Fisher确切概率法,并计算OR值及其95%CI来评估胃癌患者的相对风险。P<0.05为差异有统计学意义。

图1 LAIR1基因3’UTR的6个SNP位点的测序图
2 结果
2.1 LAIR1基因3’UTR的多态性检测结果 两组患者LAIR1基因3’UTR的 6个SNP位点基因分布频率均符合Hardy-Weinberg平衡(均P>0.05)。胃癌组患者rs199714928位点G/A基因型及等位基因A分布频率均明显低于对照组,G/G基因型及等位基因G分布频率均明显高于对照组,差异均有统计学意义(均P<0.05),见表 2~3。胃癌组 rs1054033 与 rs6509867 位点均存在完全单倍型连锁现象,而对照组有16例存在不完全单倍型连锁现象(P<0.05),见表4。
2.2 LAIR1基因3’UTR的多态性与胃癌分化程度的关系 不同胃癌分化程度患者LAIR1基因3’UTR的多态性位点基因分布频率均符合Hardy-Weinberg平衡(均P>0.05)。胃癌低分化患者rs3826753/rs3826754位点A/A基因型及等位基因A分布频率均明显低于中高分化者,等位基因G分布频率明显高于中高分化者,差异均有统计学意义(均P<0.05),见表5~6。不同性别、肿瘤类型、TNM分期、淋巴结转移阳性率患者LAIR1基因3’UTR的SNP位点基因分布频率比较,差异均无统计学意义(均P>0.05)。

表2 两组患者LAIR1基因3’UTR区6个SNP位点基因型分布频率比较[例(%)]

表3 两组患者LAIR1基因3’UTR区6个SNP位点等位基因分布频率比较[例(%)]

表4 两组患者LAIR1基因3'UTR区rs1054033与rs6509867等位基因连锁比较[例(%)]
3 讨论
LAIR1早在1997年就被克隆,同时发现它被DX26抗体识别与结合后可抑制NK细胞的细胞毒功能[3-5]。LAIR1是由287个氨基酸序列组成的Ⅰ型跨膜糖蛋白,包含了独特的膜外C2型Ig样区域和细胞质尾的2个ITIM基序[1,6]。根据分化抗原簇,LAIR1又被命名为CD305,在结构上与其他定位在染色体19q13.4上的淋巴细胞受体复合物的抑制性Ig超家族成员相似。研究发现,LAIR1基因参与抑制粒细胞-单核细胞刺激因子(GM-CSF)诱导急性髓系白血病细胞增殖,与胶原蛋白结合后强烈抑制GM-CSF介导的细胞内钙增加、磷酸化、AKT1/PKBα活化[7]。LAIR1基因的功能性配体是胶原蛋白,当羟脯氨酸存在时具有高亲和性。体外研究发现,不管是细胞系还是原发细胞中的胶原蛋白,与LAIR1基因结合均能直接抑制免疫细胞的生物学活性[8]。LAIR1基因广泛表达且其周围存在大量配体,因此必须调节LAIR1基因/胶原蛋白的相互反应,从而确保免疫系统的平衡。一方面,作用于细胞的活化信号强度由LAIR1基因/胶原蛋白的交联反应来决定信号是否被抑制;另一方面,LAIR1基因表达水平决定其与胶原蛋白反应的抑制水平。

表5 LAIR-1基因3’UTR区6个SNP位点基因型分布频率与胃癌分化程度的关系
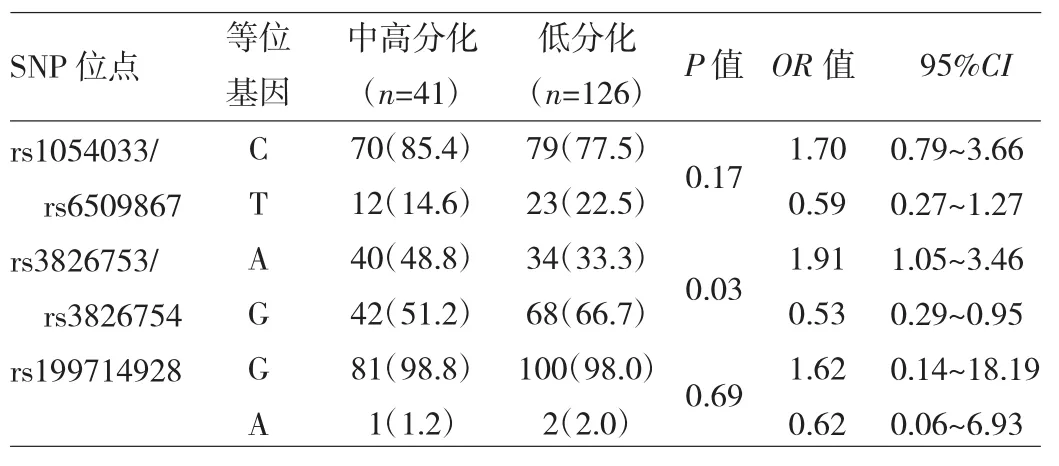
表6 LAIR-1基因3’UTR区6个SNP位点等位基因分布频率与胃癌分化程度的关系
目前关于LAIR1基因与实体瘤的关系研究较少。已有研究表明肿瘤患者血清可溶性LAIR1基因表达高于正常人,且外周血中NK细胞、CD4+及CD8+T细胞表面LAIR1基因表达明显增加[9]。研究表明,宫颈癌组织及上皮卵巢癌组织LAIR1蛋白表达均高于正常癌旁组织,且与肿瘤大小、病理分期、临床分级和淋巴结阳性数有关[10-11]。将LAIR1基因转染到不表达LAIR1基因的Me-180细胞并使其稳定表达,结果发现过度表达LAIR1蛋白会抑制细胞的增殖能力和抗凋亡能力[10]。然而,对表达LAIR1基因的HO-8910细胞进行基因敲除并使其基因表达沉默,结果发现细胞增殖能力及细胞侵袭能力反而增强[11]。
本研究对胃癌与慢性胃炎患者LAIR1基因3’UTR的多态性进行检测,结果发现胃癌患者rs199714928位点G/G基因型及等位基因G分布频率均明显高于慢性胃炎患者;但在中国西双版纳傣族人种(93例)中发现此位点全为纯化子(G/G),与此位点祖先等位基因G一致,这表明rs199714928位点部分G突变成A可能是生物进化的原因,以保护机体抵抗胃癌的发生。同时还发现胃癌患者rs1054033与rs6509867位点均存在完全的C-G和T-T单倍型连锁现象,而慢性胃炎患者存在16例突破该连锁现象的T-G单倍型。可见,rs1054033与rs6509867位点出现T-G单倍型是一种保护机体抵抗胃癌发生的现象。进一步分析发现,胃癌低分化患者rs3826753/rs3826754位点A/A基因型及等位基因A分布频率均明显低于中高分化者,等位基因G分布频率明显高于中高分化者;而不同性别、肿瘤类型、TNM分期、淋巴结转移阳性率患者LAIR1基因3’UTR的SNP位点基因分布频率比较,差异均无统计学意义。
综上所述,LAIR1基因3’UTR的rs199714928位点多态性可能是胃癌发生的易感因素;rs1054033与rs6509867位点存在不完全单倍型连锁(T-G单倍型)可能是一种保护机体抵抗胃癌发生的有利因素。
猜你喜欢
传染病信息(2022年4期)2022-11-23
河南医学研究(2022年18期)2022-09-30
世界科学技术-中医药现代化(2022年3期)2022-08-22
川北医学院学报(2022年6期)2022-06-24
昆明医科大学学报(2022年2期)2022-03-29
昆明医科大学学报(2021年6期)2021-07-31
智慧健康(2021年17期)2021-07-30
昆明医科大学学报(2021年3期)2021-07-22
中国产前诊断杂志(电子版)(2020年1期)2020-05-21
遵义医科大学学报(2020年6期)2020-02-05
