
非结构性糖在植物对干旱胁迫响应与适应中的作用
2018-11-02 03:51曹家畅崔明昆
安徽农业科学 2018年30期
周 倩,曹家畅,崔明昆,龚 明
(云南师范大学生命科学学院, 生物能源持续开发利用教育部工程研究中心, 云南省生物质能与环境生物技术重点实验室,云南昆明 650500)
近年来,随着全球气候变化,世界各地干旱事件发生的强度、频率和持续时间不断增加,植物也越来越容易受到干旱胁迫的影响,干旱胁迫导致大量的植物生长不良甚至死亡,并且呈加剧趋势,植物体内代谢途径受其影响很大,进而影响碳水化合物的含量。碳水化合物广泛存在于自然界中,且含量十分丰富,是植物光合作用的主要产物,也是植物维持生命活动的基础物质[1-2]。按其存在形式可分为结构性碳水化合物和非结构性碳水化合物,其中非结构性碳水化合物主要包括葡萄糖、果糖、蔗糖、棉子糖、淀粉、海藻糖等,是参与植株生命代谢的重要物质[3],在植物的抗性研究中具有重要地位。
经研究发现,这些非结构性碳水化合物在植物应对干旱胁迫中发挥着重要作用。一是作为碳源缓冲物质,暂时维持植物的生长和代谢;二是当作信号分子,参与调节植物体一系列代谢活动,对植物的基因表达、逆境响应、生长发育进行调控[4];三是作为渗透调节物质,保持膨压而维持正常的代谢活动[5];四是充当渗透保护剂,维持细胞质膜、蛋白质、核酸等生物大分子空间结构和功能活性[6-7]。
1 干旱胁迫对植物的影响
干旱因为长期缺少水源,使植物吸收的水分远远低于植物体本身蒸腾散失的水分,致使植物体严重缺水,生理生化和形态都发生不良变化的现象。在干旱和半干旱地区,干旱胁迫会直接影响植物的生长和生存。目前,为了适应水分供应不足,植物已经发展形成不同的生理机制,包括渗透调节、气孔开闭机制以及根部形态特征的修饰等[8-9]。维持叶片水分状况和高效光合作用对干旱条件下的可持续生长是非常重要的。植物与环境的长期相互作用,形成独有的抗旱系统。植物现有3种不同的抗旱机理,一是逃避干旱,一般发生在生命周期短的植物中,它们在干旱胁迫前就完成生命周期,以逃避干旱的危害;二是抵御干旱,植物通过大量吸收水分并尽量降低水分流失来拮抗干旱;三是忍受干旱,即在水分严重不足的情况下仍然存活,这种在低等植物中比较常见[10]。
1.1形态结构的变化干旱胁迫下,植物在形态结构上会有明显的变化,其中叶片最为敏感,在轻度的水分胁迫下叶片的生长都会受到抑制,另外植物的茎秆会变得矮小。已有研究表明,长白山地区的红松、槭树等树种为适应当地的干旱环境,反应出降低叶片面积、根长、侧根等生理指标[11]。叶具表皮毛,避免强光的照射,减少水分蒸腾;叶片表皮外壁的角质层增厚,减少水分的散失。除此之外,植物的气孔会下陷、关闭,叶片蜡质也会增加等[12]。
此外,植物的根系会变大,不断往下延伸变得密集,这是因为根系是连接土壤-植株-大气间水循环过程中的重要步骤,是植株吸收水分的主要器官,能够感知土壤环境水分的改变,从而调节自身体内的水分运输和吸收[13]。同时木质部的导管直径和数目也会增加,减少阻力提高运输水分的效率等[14];茎中机械组织发达,木质化高,机械支持增强,输导组织得到保护,同时防止大风伤害幼茎,这些都是长期缺水形成的适应性特征[15]。
1.2对光合作用的影响植物光合作用和生长(生物量生产)是受干旱影响的主要过程[16]。干旱胁迫下光合作用会减弱,由于水分亏缺,叶片含水量会相应下降,随之叶片表面保卫细胞失水,细胞膨压降低,结果造成气孔关闭,减少失水;然而植物正常进行光合作用所需的CO2不能保证,造成CO2亏缺,导致光合速率减弱[17]。
水分胁迫下,导致光合作用减弱的2个影响因素分别是气孔限制和非气孔限制[18-19]。气孔限制间接影响光合作用,因为水分缺失造成气孔关闭,气孔对CO2的导性降低,导致光合速率降低;而非气孔限制直接影响光合作用,通过影响叶绿体的显微结构、光化学反应的活力、RuBP和PEP等酶的活性来减弱光合作用。在水分胁迫的早期,引起光合作用下降的主要是气孔限制,当胁迫加重到中度以上后,非气孔因素成为主要因素。张仁和等[20]研究表明,玉米在重度干旱下,光合作用下降是受非气孔限制的。不论是否是气孔因素引起的光合作用减弱,最终都是降低碳同化速率,影响光合产物的积累。
1.3对渗透调节的影响渗透调节是植物适应干旱胁迫的重要生理机制。研究表明,干旱胁迫能够促进植物体内合成糖醇类、氨基酸及其衍生物如脯氨酸、甘氨酸、甜菜碱和多胺等低分子量的有机溶质, 且耐旱性较强的品种较抗性弱的品种上升幅度大[21]。例如, 干旱胁迫下香橙生长受到的抑制较小, 这与其叶片脯氨酸含量大幅积累有关[22]。
渗透调节物质主要有2类,一是从外界环境中进入细胞内的无机离子,主要以K+为主,可维持体内水分平衡,一些盐生或喜盐植物主要靠无机离子的积累进行渗透调节。例如柠条在受到干旱胁迫时,K+、可溶性糖、脯氨酸的含量增加,在柠条遭受干旱时有助于细胞或组织免遭脱水造成的伤害[23]。二是细胞合成的有机溶质,主要以可溶性糖(蔗糖、葡萄糖、果糖、海藻糖、棉子糖等)甜菜碱和脯氨酸为主,可调节细胞质的渗透势,维持正常的生理活动,并对蛋白质和生物膜起保护作用,从而保证细胞结构和功能的相对稳定。在干旱胁迫下,植物体内甜菜碱会积累,是由于植物体内的甜菜碱醛脱氢酶活性升高,从而增强植物的抗旱性[24]。
渗透调节是植物忍耐长期干旱过程中的一种保命机制,只是为了使植物在极端干旱的环境中存活下来,等待水分状况的改善。
2 非结构性碳水化合物代谢
2.1蔗糖代谢大多数绿色植物光合作用的终产物之一就是蔗糖,作为主要的可溶性糖,是植物体内可溶性糖的主要运输形式,对于植物生长起关键作用,面对干旱胁迫,蔗糖代谢中的非结构性糖具有重要作用,一是作为渗透调节物质,提高植物细胞的渗透势[25]。二是充当信号物质,调节转运蛋白,诱导植物体内抗性基因的表达[26]。三是作为抗氧化剂发挥作用[27]。同时,蔗糖还能影响调控花的诱导、组织分化、种子发育等过程[28-30]。
蔗糖的代谢途径是以磷酸二羟丙酮(Dihydroxyacetone phosphate,DHAP)为底物先合成磷酸甘油醛(Glyceraldehyde 3-phosphate,GAP),在醛缩酶催化下形成果糖-1,6-二磷酸,再由果糖-1,6-二磷酸酯酶(FBPase)水解形成F-6P;F-6P在磷酸葡萄糖异构酶和变位酶作用下,形成G-6P和G-1P,此3种磷酸己糖处于动态平衡状态,然后UDPG焦磷酸化酶催化G-1P和UTP合成葡萄糖供体UDPG;蔗糖磷酸合成酶(sucrose phosphate synthase,SPS)可促进形成蔗糖,而蔗糖合成酶(sucrose synthase,SS)主要是催化蔗糖分解生成果糖和UDPG,最终蔗糖在转化酶(invertase,InV)作用下分解生成果糖和葡萄糖。
蔗糖磷酸合酶(SPS)是蔗糖生物合成中的关键限速酶[31]。其是一种低丰度蛋白,含量低,且不稳定,一般与蔗糖磷酸酯酶以复合体的形式存在。蔗糖合成酶(SS)是植物细胞中蔗糖代谢的关键酶,它的催化反应是可逆的,优先将蔗糖和UDP转化为果糖和UDP-葡萄糖,UDP-葡萄糖(UDPG)是核苷酸糖在许多糖基化反应中作为葡萄糖残基的供体,并且对于蔗糖的胞质形成和参与细胞壁纤维素的合成是必需的。转化酶(InV)又称β呋喃果糖苷酶,是能够将蔗糖分解为葡萄糖和果糖的酶。它调控糖的积累和流向,是非常重要的酶之一。
2.2淀粉代谢淀粉是植物中主要的碳水化合物储存库。它是由葡萄糖残基组成的简单分子,它们通过a-1,4键与偶联的a-1,6分支相互连接,形成渗透惰性、半结晶和致密颗粒[32]。淀粉最常见的储存器官如根、根茎、块茎、茎和种子,不仅代表人类饮食中碳水化合物的主要来源,而且还为工业生产提供原料[33]。在储存器官中,淀粉由运输的蔗糖合成,可以在生长季节中储存,也可经过多年时间储存,促进再生长或幼苗生长。
淀粉代谢在叶中最为清楚(图1),其中淀粉在白天被合成,并在第二夜被动员,以确保在不可能进行光合作用时稳定地供应碳和能量。在光合作用过程中,淀粉从过量的糖质中产生,和淀粉代谢有关的酶包括ADP-葡萄糖焦磷酸化酶(ADP-glucose pyrophosphorylase polypetide,AGPase)、淀粉合成酶(starch synthase,SSS)、分支酶(starch branching enzyme,SBE)和脱支酶(starch debranching enzyme,DBE)、β-淀粉酶(β-amylase,BMY)等。其是由6-磷酸果糖(fructose 6-phosphate,F6P)转变成6-磷酸葡萄糖(glucose 6-phosphate,G6P)和1-磷酸葡萄糖(glucose 1-phosphate,G1P)。G1P在AGPase作用下形成腺苷二磷酸葡萄糖(adenosine diphosphate glucose,ADPG)之后, 在SSS、SBE和DBE的作用下合成直链淀粉和支链淀粉[34];在夜间,通过葡聚糖-水双激酶(glucan,water dikinase,GWD)和磷酸葡聚糖-水双激酶(phosphoglucan, water dikinase,PWD)磷酸化淀粉颗粒刺激淀粉降解,β-淀粉酶可将葡聚糖转化成麦芽糖,在细胞溶质中,麦芽糖转化成葡萄糖,随后形成果糖和蔗糖[35-36]。
淀粉由排列成渗透惰性颗粒的葡萄糖聚合物组成,它在大多数植物中作为主要的碳水化合物储存库,可以被快速调动,以提供可溶性糖,并且它的新陈代谢对环境的变化非常敏感。淀粉作为调节植物对非生物胁迫如水分亏缺、高盐度或极端温度反应的关键分子。在这些具有挑战性的环境条件下,当光合作用可能受到限制时,植物通常会将淀粉再生,以提供能量和碳。释放的糖和其他衍生的代谢物支持植物在胁迫下的生长,并且作为渗透保护剂和相容性溶质起作用,以减轻压力的负面影响[37]。糖也可以作为信号分子,与ABA依赖性信号通路串扰以激活应激反应级联中的下游组分[38]。另外在控制水和CO2与环境交换的气孔上的保护细胞中,淀粉在光照下会非常迅速地降解,有助于产生有机酸和糖,以增加保卫细胞的膨胀和促进气孔开放[39]。

图1 淀粉代谢途径[40]Fig.1 Pathway of starch metabolism
2.3棉子糖代谢棉子糖系列寡糖(raffinose family oligosaccha-rides,RFOs)是由一系列半乳糖基转移到半乳糖基受体上,并以α-1,6 半乳糖苷键相连而成。棉子糖、水苏糖和毛蕊花糖等是主要可溶性碳水化合物。其中棉子糖是家族中的最小成员,在植物的叶、根、种子中都有分布[41]。
棉子糖在植物体内是以葡萄糖和半乳糖为底物生成UDP-半乳糖,肌醇半乳糖苷合酶(Galaetinol synthase,GS)催化由UDP-半乳糖和肌醇合成肌醇半乳糖苷(Galactinol)。随后肌醇半乳糖苷在棉子糖合酶(raffinose synthase, RS)的作用下与蔗糖反应生成棉子糖,而后在水苏糖合酶作用下生成水苏糖,最后通过α-半乳糖苷酶作用下继续生成半乳糖和葡萄糖(图2)。
研究表明棉子糖系列寡糖作为渗透调节物质,在植物面对非生物胁迫时能够稳定光合系统,作为保护剂保护细胞[42-43]。此外,棉子糖系列寡糖还具有清除自由基的功能[44];而且还被公认参与种子的脱水性和贮藏性[45]。虽然RFOs途径在响应应激耐受方面的作用尚未被完全了解,但植物体内RFOs合成相关基因在胁迫应答中的作用越来越被重视。
2.4海藻糖代谢海藻糖(trehalose)是一种非还原性二糖,在自然界中广泛分布于动植物及细菌、真菌体内,作为应激保护剂分子对多种生物活性物质具有非特异性保护作用。大量研究表明在海藻糖丰富的生物体中,其不但可作为碳库调节生长发育,还可充当渗透保护剂维持细胞质膜、蛋白质、核酸等生物大分子空间结构和功能活性[46-47],同时还有助于降低高温、渗透、脱水应激和有毒化学物质等多种胁迫造成的损伤,提高植物的耐受性[48]。
海藻糖和蔗糖生物合成途径具有共同的特征,但其也具有与它们在细菌、真菌、藻类、植物、昆虫和哺乳动物等生物中的存在和作用相伴随的不同性质和调节[49]。与葡萄糖和蔗糖相比,不仅生物合成不同,而且海藻糖的分子性质都不同。海藻糖具有最高的玻璃化转变温度(所有二糖中最高),高度黏稠,在极端温度下保持玻璃状结构,并在干燥和冷冻条件下覆盖生物分子,从而增加这些生物分子的稳定性[48]。此外一种糖信号分子调节碳水化合物的分配,并与其他糖类的代谢途径相互作用[50]。
在植物体内海藻糖合成途径主要是OtsA-OtsB途径,以葡萄糖为底物,通过海藻糖-6-磷酸合成酶(Trehalose-6-phosphate synthase,TPS)将尿苷二磷酸葡萄糖与葡萄糖-6-磷酸结合,从而形成海藻糖6-磷酸;海藻糖-6-磷酸磷酸酶(Trehalose-6-phosphate phosphatase,TPP)催化海藻糖6-磷酸去磷酸化合成海藻糖,海藻糖最终在海藻糖酶(Trehalase,THase)作用下分解为葡萄糖[51],该途径广泛存在于生物体内(图3)。
最近的研究强调海藻糖途径在胁迫耐受中的作用,作为渗透保护剂或信号传导分子。但迄今为止,还没有记录海藻糖途径在植物胁迫耐性中的具体作用或作用模式,因为当作为渗透保护剂分子时,积累的水平通常较低,并且与该途径有关的基因的过表达可能导致不希望的特征。故海藻糖途径在提供植物抗逆性方面的确切机制仍不清楚,有待深入研究。
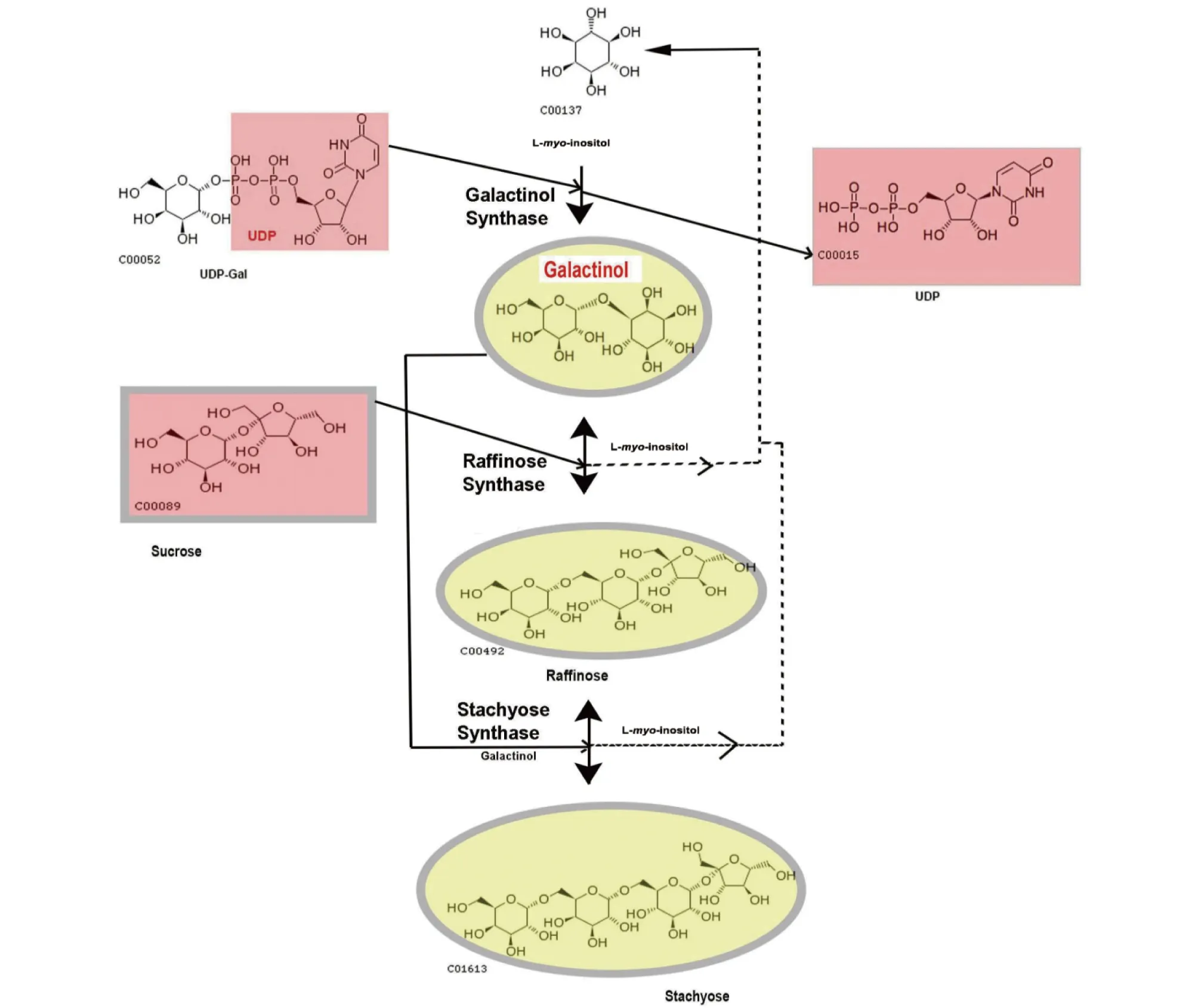
图2 RFOs合成途径[52]Fig.2 Synthesis pathway of RFOs

图3 海藻糖代谢[53]Fig.3 Metabolism of trehalose
3 结语
随着全球气候不断变暖,干旱的情况越来越突出,目前地球上1/3土地已是干旱或者半干旱,水资源的缺乏已影响农作物和森林的保护,干旱对植物的非生物胁迫占首位。植物在进化过程中已形成多种响应和适应干旱的方式,通过形态、生理、代谢和细胞水平上的多种变化来抵御干旱。糖是小分子化合物,在植物生长发育的过程中发挥着重要的作用,而且植物体内糖类的合成与分解会影响细胞的渗透势,调控细胞的吸水能力或保水能力,在植物适应干旱环境中起重要作用。随着植物抗逆性机理研究的逐渐深入,发现渗透调节是各种逆境胁迫中的共同作用,非结构性糖作为一种重要的逆境调节物质,在保护植物细胞膜功能性、碳源定向运输、ROS清除、稳定酶分子、调节细胞多空间渗透势等方面起重要的作用。
猜你喜欢
军事文摘(2023年18期)2023-10-31
科学(2022年4期)2022-10-25
干旱地区农业研究(2022年1期)2022-01-28
装备制造技术(2019年12期)2019-12-25
小学生必读(低年级版)(2019年5期)2019-08-30
Coco薇(2016年5期)2016-06-03
考试周刊(2016年6期)2016-03-11
中国铸造装备与技术(2015年5期)2015-12-10
少儿科学周刊·儿童版(2015年10期)2015-11-07
