
不同品种硫肥对苋菜镉累积的影响
2018-10-29 08:14王秀斌迟克宇霍文敏范洪黎
农业环境科学学报 2018年10期
邹 茸,王秀斌,迟克宇,霍文敏,2,王 丽,范洪黎*
(1.中国农业科学院农业资源与农业区划研究所/农业部植物营养与肥料重点开放实验室,北京 100081;2.中国地质大学,北京100083;3.北京建工环境修复股份有限公司,北京 100015)
大量重金属随工业废水和废渣等进入土壤,导致农田土壤重金属污染呈日益严重的趋势[1-2]。重金属在“土壤-植物”体系中富集、迁移和转化[3],毒害农作物,造成植物生长受阻和减产;同时,植物中累积的重金属又通过食物链等在人体蓄积,危害人体健康[4]。土壤Cd的移动性强、毒性高、污染面积大。近年来研究发现,我国湘江区域Cd的潜在生态风险很高[5],农田土壤和农作物已普遍受到了一定程度的重金属污染[6-7]。同时,植物修复由于具有对土壤理化性质影响小、成本低、效果好等优点,日益成为污染土壤修复的重要技术[8-10]。施肥能提高作物生物量、影响作物根系和茎叶的代谢过程,可改变土壤pH值以及携带竞争离子等,影响土壤溶液中重金属的有效性,从而影响植物对重金属的吸收与累积[11]。因此,施肥已成为提高植物修复效率的重要辅助措施之一。
硫是植物必需的营养元素之一,在植物的光合作用、呼吸以及碳水化合物代谢过程中起着重要的作用[12]。前人大量研究表明,在常规施肥的基础上增施硫肥可增加小麦和油菜的产量[13],也会促进植物体内植物螯合肽(Phytochaletins,PCs)的合成;PCs与Cd结合可降低Cd的活性,从而减轻Cd的毒害作用[14]。硫肥形态不同对植株从土壤中吸收重Cd金属离子影响也不同。硫酸铵可调节土壤pH,富含活性基团(氨基)配位体与重金属元素Zn、Cu、Fe等络合或螯合以减弱土壤中重金属的生物有效性[15]。硫酸盐的存在可提高土壤中可变电荷,改变土壤中Zn、Cd等重金属的植物吸附[16]。也有研究表明,在Pb和Zn污染土壤中添加硫磺,土壤中硫酸根含量增加,土壤pH值降低,醋酸铵浸提态的Pb和Zn含量也随施硫水平的增加而增加[17]。另外,硫磺处理下油菜吸硫量高于硫酸钾处理[18],相比单质硫,石膏硫可更好地降低土壤中Cd的生物有效性[19]。因此,选择适宜形态的肥料来控制Cd污染在土壤-植物中的迁移和累积,对土壤质量的改良和农业安全生产具有重要的应用价值[20]。Bo⁃brzecka等[21]通过连续三年的田间试验,观察到苋菜(Amaranshus mangostanus L.)的总含硫量比谷类高。目前,尽管对一些植株体内硫与Cd吸收和累积的内在联系已开展了一系列研究[22-24],但苋菜体内硫与Cd关联的研究鲜见。本文以苋菜为研究对象,研究不同特性硫肥料对酸性土壤中苋菜生长及硫和Cd吸收的影响,并且探讨不同特性硫肥的肥效差异,为选择适宜形态的硫肥来控制Cd污染和提高苋菜对Cd富集提供科学依据,丰富和发展硫和Cd植物体吸收的科学认识。
1 材料与方法
1.1 供试材料
供试土壤取自珠江三角洲地带的赤红壤和长江三角洲地带的黄棕壤,其理化性质见表1。苋菜为前期筛选的Cd超富集植物天星米(Amaranshus man⁃gostanus L.)[25]。
1.2 试验设计
采用温室盆栽试验,包括硫磺(升华硫,分析纯)、硫酸钙(CaSO4·2H2O,分析纯)、硫酸铵(分析纯)三种不同的硫源及对照(CK)共4种处理。施肥处理硫含量一致,每1.00 kg土壤含硫量为50 mg。试验中Cd的添加浓度与基肥施用量以前期苋菜对Cd污染土壤修复潜力的土培试验为依据[25],每盆装风干土0.75 kg,每1.00 kg土壤加 Cd(CdCl2·2.5H2O,分析纯)15 mg。在播种之前,Cd以CdCl2溶液形式加入,基础肥料以基肥形式加入,使用尿素(46%,每盆321.9 mg)、磷酸氢二钾(分析纯,每盆329.1 mg)与土壤充分混匀,随机设计,重复3次。放置2周后,播种,出苗一周后定植为4~5株,生长期内定期浇去离子水,保持土壤相对含水量为田间持水量的70%左右。本试验未平衡施加硫酸铵带入土壤的氮肥,因此硫酸铵处理所产生的结果是硫、氮肥共同影响的结果。
1.3 收获与测定
植株生长45 d后收获,分地上部和根部用去离子水清洗干净、105℃杀青30 min、70℃烘干。植株硫含量的测定:称取0.300 0~0.400 0 g植物样品于消煮管中,加入玻璃珠2颗和浓硝酸3 mL且加盖小漏斗过夜,再加入HClO42 mL慢慢加热至235℃消煮2 h。取下漏斗,加入HCl 1 mL,在150℃下加热20 min,冷却,用蒸馏水与缓冲液定容[26],同时做试剂空白试验。植株Cd含量测定参照食品安全国家标准GB 5009.15—2014《食品中镉的测定》:称取样品1.000 g左右,置于25 mL消煮管中,加入10 mL体积比4∶1的优级纯HNO3-HClO4混合酸,冷消化过夜。第2 d,将消煮管置于砂浴上,在165~175℃下消化,直至溶液变为无色透明,稍冷却后,分别加两次约1 mL的蒸馏水排酸,蒸发浓缩消化液至2 mL左右,用5%的HCl转移并定容至10 mL比色管,同时做试剂空白试验[27]。用ICP-AES测定其中的Cd和硫含量[26]。根据国家标准物质测定标准GBW 10020(GSB-11)《柑橘叶成分分析标准物质》:柑橘叶Cd标准值为(0.17±0.02)mg·kg-1,本试验中的柑橘叶Cd测定值为0.18 mg·kg-1符合GBW 10020(GSB-11)要求。用SAS 9.4软件对试验数据进行方差分析和多重比较[28]。根据生物富集系数和转运系数计算方法,计算植株生物富集量系数和转运量系数来反映其吸收和累积重金属Cd的能力[29]。
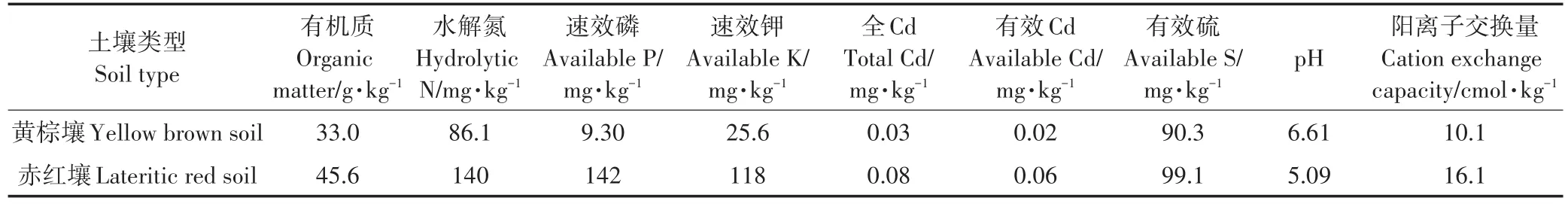
表1 土壤基本理化性质Table 1 Physical and chemical properties of soil
生物富集系数=植株地上部Cd含量(mg·kg-1)/土壤中Cd含量(mg·kg-1)
转运系数=植株地上部Cd含量(mg·kg-1)/植株根部Cd含量(mg·kg-1)
2 结果与分析
2.1 不同硫源对苋菜生物量的影响
在氮磷钾供应充足的基础上,硫供应水平提高可促进植株生长、增产,但增施硫肥的形态不同增产效果也不同。由图1可知,在黄棕壤中,硫酸铵、硫酸钙处理的地上部和根部生物量显著高于对照和硫磺处理的生物量,并且硫酸铵处理增幅最大;与对照相比,地上部、根部干质量分别增加了36.4%和37.5%,硫磺处理植株干质量增量最小且地上部干质量与对照无显著差异。在赤红壤中,不同种类的硫肥处理地上部、根部干质量差异显著,且硫肥处理后地上部与根部生物量变化趋势相同:硫酸铵处理>硫酸钙处理>硫磺处理>对照。同种硫肥在两种不同类型土壤中施用,皆表现为黄棕壤上苋菜的生物量较大。从结果可以看出,硫酸铵处理对于苋菜植株生长的促进作用最为明显,硫磺处理对于苋菜植株生长的促进作用最小。
2.2 不同硫源对苋菜吸收和累积Cd的影响
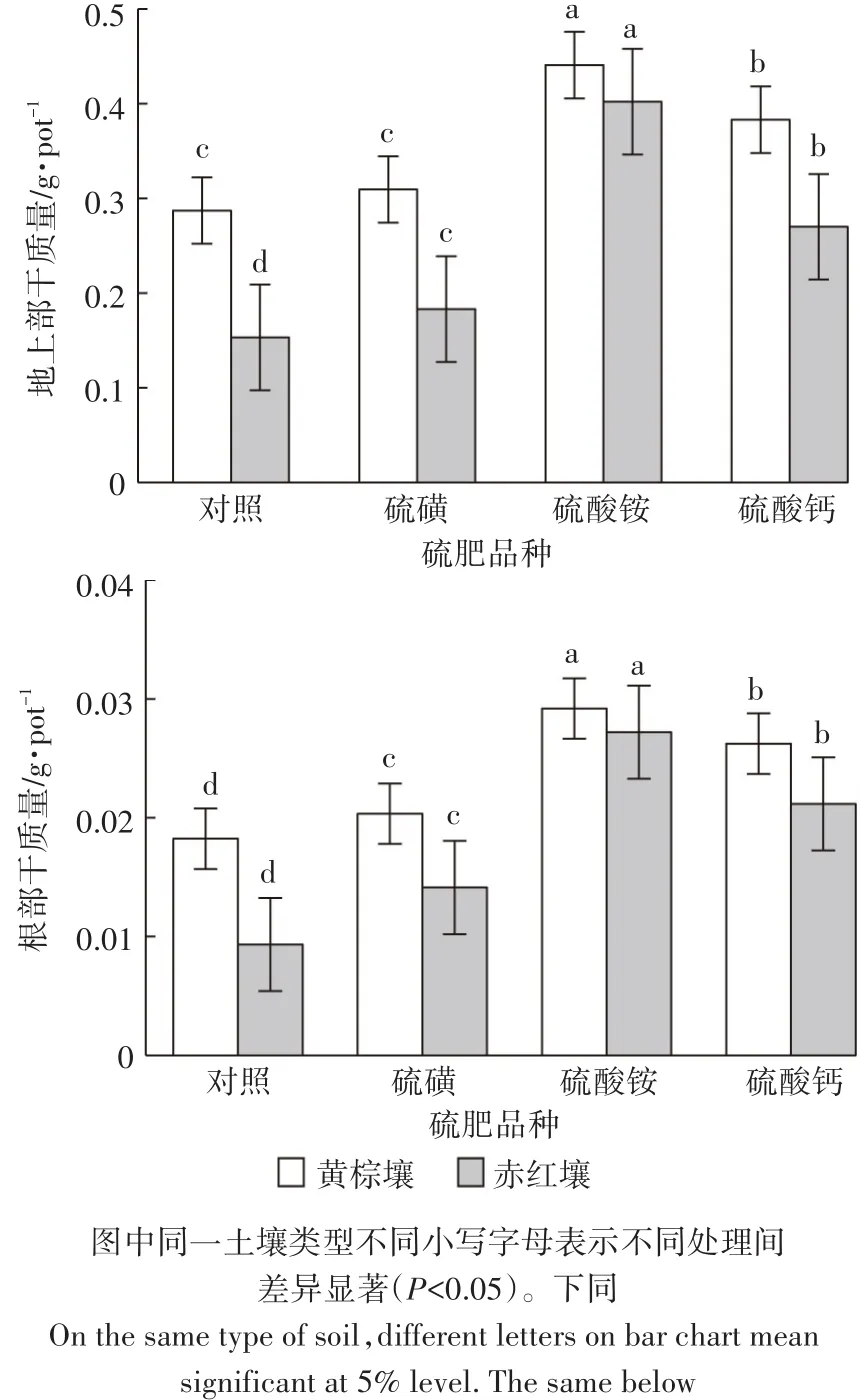
图1 不同硫肥品种对苋菜生物量的影响Figure 1 Effects of different sulfuric fertilizers on the biomass of Amaranshus mangostanus L.
增施硫肥会促进植株的生长,也会影响土壤理化性质、改变土壤Cd的生物有效性,从而影响苋菜对Cd的吸收和转移。由表2可见,Cd污染的两种土壤施加硫肥,各处理间苋菜植株Cd含量和Cd累积不同。黄棕壤中,不同硫肥皆促进了苋菜对土壤Cd的吸收和累积。硫酸铵处理的地上部、根部Cd含量最大,比对照分别高约60%、130%,与其他处理相比,硫酸铵处理苋菜地上部、根部Cd累积量最高,地上部和根部分别是对照的2.45、3.68倍。相对硫酸铵与硫酸钙处理,施用硫磺苋菜地上部、根部的Cd含量及其Cd累积量最小。比较苋菜植株对重金属Cd的吸收和转运能力,可发现生物富集量系数大小为:硫磺>硫酸钙>对照>硫酸铵;植株转运量系数高低顺序为:硫磺>对照>硫酸钙>硫酸铵。硫磺处理下苋菜的富集、转运能力最高。
赤红壤中,各个硫肥处理地上部Cd含量显著高于对照,硫酸铵处理含量最高,比对照高29%;其中硫磺与硫酸钙处理间无显著差异,Cd累积量的变化次序大致为:硫酸铵>硫酸钙>硫磺>对照,硫酸铵处理的地上部Cd累积量是对照的3.40倍。硫酸铵处理的根部含Cd量和累积Cd量最大,分别是对照的2.24倍和6.45倍,且与其他三个处理差异显著,而相对硫酸铵与硫酸钙处理,施用硫磺苋菜地上部、根部的Cd含量及其Cd累积量最小。其中,硫酸铵处理的生物富集量系数最大,而硫磺处理的转运量系数最大。
结果显示,Cd污染的黄棕壤中,硫酸铵处理能使苋菜最大限度地吸收和累积土壤中的Cd;而硫磺处理利于苋菜植株根部更好地吸收和转移Cd2+,减轻Cd对植株根系的毒害作用。在Cd污染的赤红壤中,硫酸铵处理和硫磺处理均有利于Cd从苋菜植株根部向地上部转移。由苋菜地上部和根部Cd累积量可知,施加同一硫肥时,黄棕壤上苋菜Cd累积量大于赤红壤。因此,不同类型土壤的理化性状不同,其硫肥处理的效果也存在很大差异。
2.3 不同硫源对苋菜吸收和累积硫的影响
硫肥的增施可促进植株对硫的吸收和同化。由图2可知,黄棕壤中,各个硫肥处理苋菜地上部含硫量皆高于对照,且硫酸铵处理含量最高,为对照的1.11倍;根系含硫量,硫酸铵、硫酸钙和硫磺处理均高于对照,分别较对照高约18%、62%和36%。结果表明,施加不同种类的硫肥均促进了苋菜根部对硫的吸收与累积;由于硫在植物体内的移动性有限,地上部硫含量变化不及根部大。赤红壤中,硫酸铵、硫磺和硫酸钙处理的地上部硫含量分别是对照的1.18、1.23倍和1.09倍,根系硫含量各处理间存在显著差异,硫酸铵>硫酸钙>硫磺>对照。因此,施加硫肥皆可增加苋菜的硫含量,以硫酸铵效果最好。
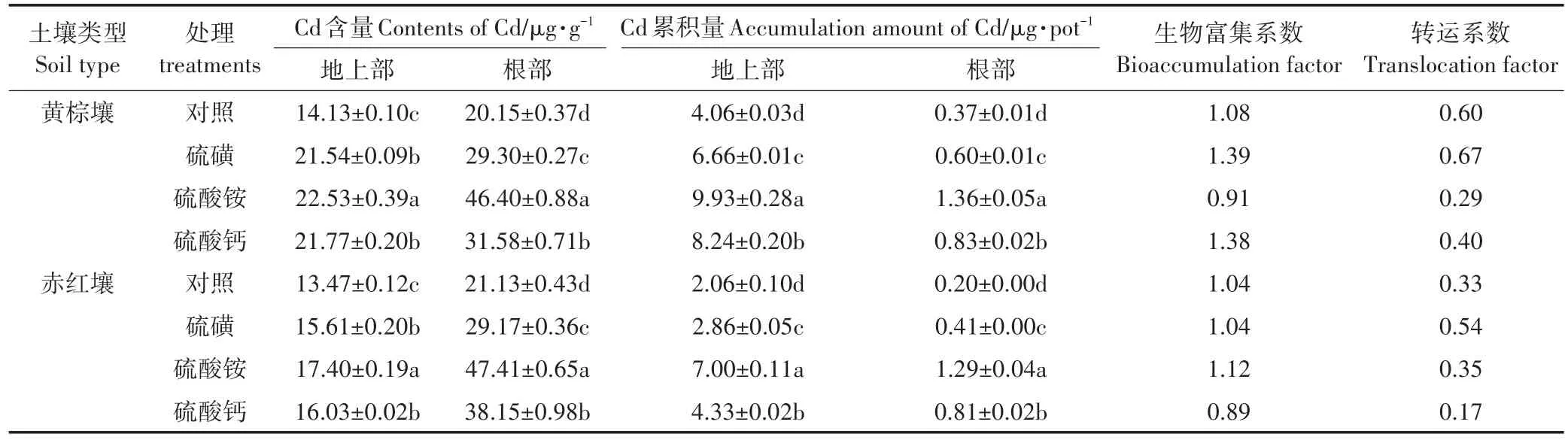
表2 不同硫肥对苋菜植株体内Cd浓度和累积量的影响Table 2 Effects of different forms of sulphur on accumulation of Cd and content of Cd in Amaranshus mangostanus L.

图2 不同硫肥对植株吸收和累积硫的影响Figure 2 Effects of different forms of sulphur on uptake of S and content of S in Amaranshus mangostanus L.
图2可见,黄棕壤中,苋菜植株地上部总硫量为硫酸铵>硫酸钙>硫磺>对照,硫肥处理根部硫累积量皆显著高于对照,其中硫酸铵处理是对照的1.62倍;赤红壤中,硫酸钙、硫酸铵、硫磺三个处理地上部硫累积量显著高于对照,分别是对照的1.41、3.22倍和2.07倍。黄棕壤中,增施硫肥苋菜根部硫累积量增加且各处理大小顺序为:硫酸铵>硫酸钙>硫磺>对照;赤红壤中苋菜根部硫总量变化趋势与黄棕壤相同,且硫酸铵、硫酸钙和硫磺处理分别是对照的3.97、2.85倍和1.76倍。因此,在两种不同类型土壤上施加不同种类硫肥都能促进苋菜地上部对硫积累,且两种土壤上根系与地上部硫累积量变化趋势相同。结果表明,在三种硫肥处理中,硫酸铵促进植株吸收和累积硫的效果较好,而硫磺的效果较差。
3 讨论
3.1 不同硫肥对苋菜生物量及硫吸收和累积的影响
土壤中的硫主要是以硫酸根离子的形态,以质流的方式被植物主动吸收,增施适量硫肥可促进植株生长。赵玉霞等[30]通过大田试验研究发现,增施硫肥冬小麦产量增加,全生育期内各器官的吸硫量随施硫量的增加而提高,有显著增产效果。外施硫肥也能明显缓解Cd胁迫对小白菜叶片光合作用的抑制,促进了小白菜的生长[31]。植物在Cd污染的土壤中生长的能力与硫还原同化过程密切相关。Cd胁迫下高亲和与低亲和硫转运体基因在根中过量表达,增加植物体对SO2-4的吸收量,合成较多的含硫化合物用以抵抗Cd毒害[32]。另外,增施硫肥硫素同化和谷胱甘肽代谢增强可降低Cd易位[33],清除过量Cd引发的活性氧,同时促进植物体内非蛋白巯基(NPT)、GSH和PCs等含硫化合物合成[34],Cd与PCs结合后,被运输至液泡,形成高分子量的复合物,从而使细胞处于无毒化状态[35],抑制Cd向其他细胞器中转运,保证细胞器功能正常运转。也有研究发现,Cd对油菜叶片内活性氧清除酶活力的诱导提高并非是抵抗Cd胁迫的关键途径,而与硫代谢有关的对Cd的络合及固定作用减轻对叶片的伤害才是其重要的抗Cd机制[36]。本研究结果表明,增施硫肥皆促进了苋菜对硫的吸收和累积,提高了苋菜的生物量,不同硫肥效果不同。硫酸铵促进作用最为明显,硫酸铵不仅可被根系直接吸收,而且也为苋菜提供了一定量的氮肥,促进了苋菜对氮和硫的吸收;硫磺不能直接被植株吸收,施入土壤后需氧化为硫酸根才能被根系吸收,受到土壤微生物、通气状况、土壤温度、湿度等多种复杂因素的影响,因此,硫磺处理苋菜硫含量及累积量较少,苋菜生物量较小。另外,同种硫肥施在两种土壤上时黄棕壤苋菜生物量皆高于赤红壤,这可能是苋菜对硫的吸收和累积与土壤的物理化学性质有密切关系,还需要进一步的研究探索。
3.2 外源硫肥对苋菜吸收和累积Cd的影响
本试验中,在有效硫含量大约90 mg·kg-1Cd污染的两种酸性土壤上,施入三种不同特性硫肥,苋菜的Cd累积量也随之增加。前人研究发现,缺硫阻碍了Cd向地上部运输,导致地上部Cd的含量明显降低[37];李会合等[38]研究表明,用含 1.5~2.25 mmol·L-1硫的营养液培养东南景天可显著提高其根、茎和叶的Cd含量、累积量及整株累积总量。土壤pH对Cd形态和有效性影响大,硫酸铵为生理酸性肥料,植物吸收后,根际土壤的pH值降低;元素硫在土壤中能被硫氧化细菌氧化成硫酸根离子且导致土壤pH下降[39],Cd的有效性皆增加。也有前人研究发现腐植酸复合肥、硫酸铵和磷酸二铵配施以及硫酸铵、磷酸二铵和硫酸钾配施2种施肥措施的表层土壤有效Cd含量低于对照[40],外源添加K2SO4会减少小麦(Triticumaestivum L.)对Cd的吸收[41],这些研究与本试验结果有差异,可能是采用的施肥方式不同、肥料形态不一致,也可能是试验作物对Cd的吸收特性不同。从苋菜地上部、根部Cd累积量看,施加同一硫肥时,黄棕壤上苋菜Cd累积量大于赤红壤。因此可推断,不同类型土壤的理化性状不同,其硫肥处理的效果也存在很大的差异。
4 结论
在黄棕壤和赤红壤上施加不同种类的硫肥,减轻Cd对植株的毒害作用并显著提高了苋菜的生物量,促进了苋菜对硫的吸收和累积。施用不同种类的硫肥,苋菜对Cd的吸收和累积存在一定的差异。
以50 mg·kg-1的纯硫量将不同特性硫肥施入黄棕壤、赤红壤中,均能显著提高苋菜地上部和根部的Cd累积量,其中以施加硫酸铵效果最佳;在黄棕壤中施加硫磺,苋菜生物富集系数及转运系数最大,在赤红壤中施加硫酸铵,苋菜的富集系数最大,施加硫磺,苋菜的转运系数最大,施加硫磺更利于苋菜植株根部更好地吸收和转移Cd离子。在一定土壤Cd浓度范围内,施用硫酸铵可提高苋菜Cd累积量,且成本低、效果好、易操作,适宜推广应用。
猜你喜欢
中国化肥信息(2022年6期)2023-01-06
中国化肥信息(2022年3期)2023-01-05
应用化工(2022年5期)2022-07-11
中国化肥信息(2022年4期)2022-06-07
课外语文·中(2022年1期)2022-02-16
无机盐工业(2022年1期)2022-01-19
老友(2021年6期)2021-07-01
百科知识(2021年2期)2021-02-24
中国化肥信息(2020年3期)2021-01-07
中华养生保健(2020年3期)2020-11-16
