
E-cadherin、TGF-β1和Twist1在结直肠癌变过程中的表达及意义
2018-10-12 10:53薛敬玲
数理医药学杂志 2018年10期
温 辉 薛敬玲
(武汉大学基础医学院 武汉 430071)
结肠直肠癌,一种最常见的起源于上皮细胞的上皮癌,是工业化国家癌症相关死亡率的主要原因,在西方国家结肠癌占恶性肿瘤死因的第3位[1]。在我国,据统计结直肠癌发病率为29.44/105,死亡率为14.23/105,且其发病率与死亡率呈逐步上升的趋势[2]。Morson等研究者提出了腺瘤-癌演变学说[3],认为结直肠癌的发生起源于腺瘤的癌变,结直肠腺瘤属癌前期病变,上皮细胞出现异常增殖,逐渐形成腺瘤性息肉组织,最终形成恶性肿瘤组织。近年来研究发现肿瘤浸润转移及局部复发是导致其治疗失败和患者死亡的主要原因,上皮-间质转化(epithelial-mesenchymal transition, EMT)是许多肿瘤侵袭和转移早期的一个重要的过程[4],其中多个转录因子参与 EMT 的调节。
Twist1是一种碱性螺旋状的转录因子,在恶性肿瘤疾病发生发展中发挥作用,还可明显促进肿瘤出现侵袭和转移等病理现象[5]。转化生长因子β1(transforming growth factor β1,TGF-β1)则是具有多种生理学功能的多肽类生长因子,可有效抑制细胞生长,促进细胞间基质的形成,与肿瘤疾病发生、发展存在密切的联系[6]。上皮性钙黏蛋白(E-cadherin)是一种钙黏蛋白,其表达水平降低可导致上皮细胞之间的黏附能力明显丧失、极性消失,使得肿瘤细胞迁移能力显著性增强,最终导致肿瘤出现侵袭和转移等[7]。本研究拟检测E-cadherin、TGF-β1和Twist1在结直肠癌组织中的表达,并探讨其临床意义。
1 资料与方法
1.1 临床资料
选择我院2016年1月~2017年12月间手术切除、病理证实为结直肠癌(腺癌) 的标本70例,其中男性45例,女性25例,年龄18~80 岁,组织病理学确定病理类型、分化程度、浸润深度、微浸润情况及有无局部淋巴结转移。组织分化程度:高、中、低分化分别36、20、14例;浸润深度: 至或者穿透浆膜层的42 例,到达肌层的28 例;有局部淋巴结转移的26 例,无转移的44例;30 例癌旁正常组织取自距肿瘤边缘5 cm 以上黏膜;结直肠高级别上皮内瘤变60例,结直肠低级别上皮内瘤变60例。
1.2 研究方法
采用免疫组织化学方法检测各组E-cadherin、TGF-β1和Twist1表达水平,将所有切除后新鲜标本固定在10%中性缓冲甲醛固定液中,取出标本,经脱水、透明、浸蜡、包埋等步骤后,切片,片厚3μm,测定时进行二甲苯脱蜡及梯度乙醇水化,3%过氧化氢浸泡10min灭活内源性过氧化物酶,柠檬酸缓冲液( pH 6.0±0.1)高温修复抗原;使用一抗分别为:兔抗人TGF-β1单克隆抗体(购自武汉博士德生物技术有限公司,工作液浓度为1∶200) 、兔抗人Twist1多克隆抗体(购自上海爱必信生物技术有限公司,工作液浓度为1∶1600) 、鼠抗人E-cadherin单克隆抗体(购自福州迈新生物有限公司, 即用型),使用二抗是即用型免疫组化EliVision检测试剂盒(购自福州迈新生物有限公司)。采用EnVision二步法:滴加一抗,4℃冰箱孵育过夜;滴加二抗,37°C孵育30min;各步骤间均以0.01 mol /L PBS 缓冲液冲洗,严格按照说明控制时间和温度;DAB显色、苏木素轻微衬染、脱水、透明、中性树胶封片,用PBS代替一抗作阴性对照,用已知阳性切片作为阳性对照,显微镜观察。
1.3 判断标准
E-cadherin和TGF-β1按Shimizu[8]方法对每张切片阳性细胞的着色程度进行分析,分为着色、淡黄色、棕黄色、棕褐色,分别计为0、1、2、3分,着色阳性面积按无着色、着色面积<1/3、1/3~2/3、>2/3分别计0、1、2和3分,根据着色程度和着色面积积分之和判断最终实验结果:≥3分为阳性,<3分为阴性,随机选择5个视野,取其平均值。Twist1主要表达于细胞核,根据阳性细胞数的比例将染色结果分为阳性与阴性[9]:即阳性细胞数0~10%为阴性,阳性细胞数>10%为阳性,所有切片由两名具有丰富阅片经验的病理医师以双盲法予以判读。
1.4 统计学方法
采用SPSS16.0软件包对全部资料予以分析处理,计数资料采用χ2检验,相关性采用非参数Spearman等级进行相关分析,以P<0.05表示差异有统计学意义。
2 结果
2.1 各组E-cadherin、TGF-β1和Twist1阳性率比较
E-cadherin、TGF-β1和Twist1蛋白在不同组织中的表达结果见图1。免疫组化结果显示了3种蛋白在癌组织中的定位:E-cadherin在正常细胞主要定位于细胞膜,而在肿瘤细胞定位于细胞膜和细胞质, TGF-β1定位于癌细胞的细胞质中,而Twist1表达定位主要存在于癌细胞细胞核中,部分细胞质也有表达。
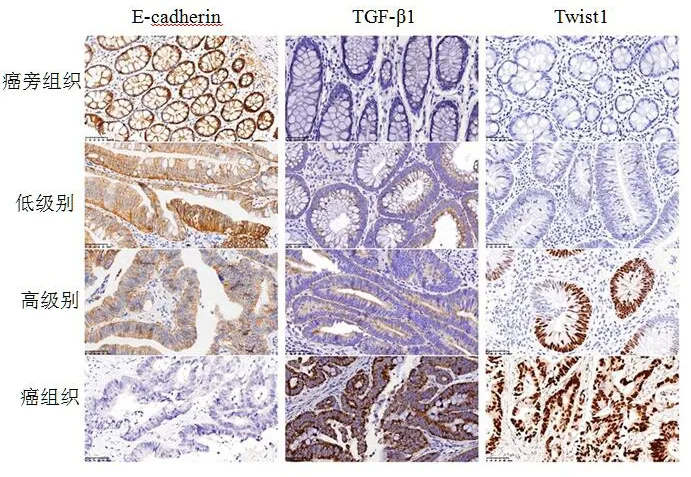
图1 结直肠癌组织、高级别上皮内瘤变组织、低级别上皮内瘤变组织、癌旁组织中E-cadherin、TGF-β1和Twist1蛋白表达差异(免疫组化SP×200)
免疫组化结果显示各组E-cadherin、TGF-β1和Twist1阳性率比较均有显著性差异(P<0.05),且随着结直肠癌组织、高级别上皮内瘤变组织、低级别上皮内瘤变组织、癌旁组织,E-cadherin阳性率明显升高(P<0.05),而TGF-β1和Twist1阳性率明显降低(P<0.05),见表1。
2.2 结直肠癌E-cadherin、TGF-β1和Twist1表达与临床病理特征的关系
结直肠癌E-cadherin、TGF-β1和Twist1表达在分化程度、Duke分期、浸润深度、淋巴结转移等方面均存在显著性差异(P<0.05),而在性别、年龄等方面无明显差异(P>0.05),见表2。
2.3 结直肠癌E-cadherin、TGF-β1和Twist1表达之间的相关性分析
结直肠癌E-cadherin与TGF-β1、Twist1均呈负相关性关系(r=-0.632,-0.584,P<0.05),TGF-β1与Twist1呈正相关性关系(r=0.617,P<0.05)。
表1 各组E-cadherin、TGF-β1和Twist1阳性率比较(%)

组别例数Cadherin阳性率TGF-β1阳性率Twist1阳性率结直肠癌组织7042.86(30/70)85.71(60/70)80.0(56/70)高级别上皮内瘤变组织6061.67(37/60)*68.33(41/60)*63.33(38/60)*低级别上皮内瘤变组织6075.0(45/60)*#50.0(30/60)*#45.0(27/60)*#癌旁组织30 90.0(27/30)*#▲33.33(10/30)*#▲30.0(9/30)*#▲
注:*与结直肠癌组织比较,P<0.05;#癌旁组织、低级别上皮内瘤变组织与高级别上皮内瘤变组织比较,P<0.05;▲癌旁组织与低级别上皮内瘤变组织比较,P<0.05。
表2 结直肠癌E-cadherin、TGF-β1和Twist1表达与临床病理特征的关系
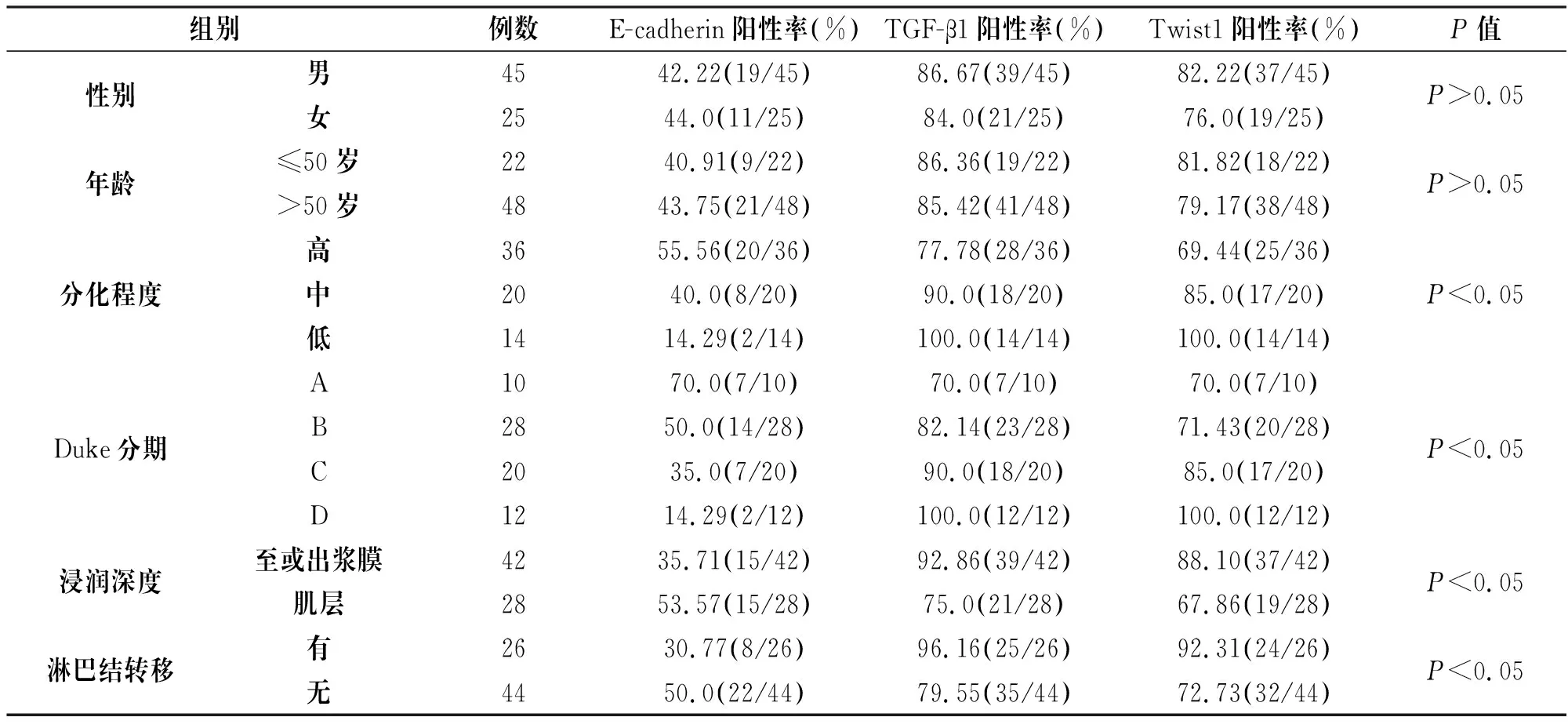
组别例数E-cadherin阳性率(%)TGF-β1阳性率(%)Twist1阳性率(%)P值性别男4542.22(19/45)86.67(39/45)82.22(37/45)女25 44.0(11/25)84.0(21/25)76.0(19/25)P>0.05年龄≤50岁2240.91(9/22)86.36(19/22)81.82(18/22)>50岁4843.75(21/48)85.42(41/48)79.17(38/48)P>0.05分化程度高3655.56(20/36)77.78(28/36)69.44(25/36)中2040.0(8/20)90.0(18/20)85.0(17/20)低1414.29(2/14)100.0(14/14)100.0(14/14)P<0.05Duke分期A1070.0(7/10)70.0(7/10)70.0(7/10)B2850.0(14/28)82.14(23/28)71.43(20/28)C2035.0(7/20)90.0(18/20)85.0(17/20)D1214.29(2/12)100.0(12/12)100.0(12/12)P<0.05浸润深度至或出浆膜4235.71(15/42)92.86(39/42)88.10(37/42)肌层2853.57(15/28)75.0(21/28)67.86(19/28)P<0.05淋巴结转移有2630.77(8/26)96.16(25/26)92.31(24/26)无4450.0(22/44)79.55(35/44)72.73(32/44)P<0.05
3 讨论
E-cadherin是钙黏蛋白家族成员之一,是一类Ca依赖的糖蛋白,在上皮细胞中起着维持细胞极性及黏附力的作用,维持正常组织和器官的结构和功能,EMT过程中会出现上皮性标记物丧失,最典型就是E-cadherin蛋白的丢失,导致上皮细胞之间的黏附力丧失、极性消失,使细胞的迁移能力增强,导致侵袭和转移[10]。Twist1为在果蝇、鼠、人等生物体内的一种碱性螺旋转录因子,主要表达于胎盘、胚胎中胚层和成人的某些中胚层来源的未分化组织。最近研究发现,Twist1具有癌基因的特征,能够抑制肿瘤细胞凋亡,参与肿瘤细胞周期的调控,并通过调解肿瘤细胞上皮-间质转化(epithelial-mesenchymal transition, EMT)等参与了恶性肿瘤的发生,并且促进肿瘤侵袭转移[11]。Valdés-Mora等[5]使用实时定量PCR技术检测了54例结直肠癌及其癌旁正常黏膜Twist1的mRNA表达,发现结直肠癌Twist1的mRNA表达显著高于正常黏膜组织表达, 并且其表达还与肿瘤淋巴结转移呈显著正相关,提示Twist1是结直肠癌演进过程中一个重要因子。TGF-β1是一类具有多种功能的多肽类生长因子,主要功能是抑制细胞生长,促进基质形成,与肿瘤发生、发展密切相关,其以自分泌或旁分泌方式产生,生物学作用多样,能调节细胞增殖、分化、形态形成及刺激细胞外基质分泌、抑制免疫反应等[12, 13]。研究认为[1],TGF-β1 作为 EMT 发生的一个主要诱导剂,与E-cadherin的转录启动序列E-box结合,抑制E-cadherin的转录,通过改变细胞形态变化、上调间质细胞标记(vimentin、fibronectin)表达及下调上皮细胞标记 E-cadherin表达来诱导癌细胞发生EMT,而Twist1作为TGF-β1 信号途径的下游信号因子,我们推测TGF-β1可能是通过转录激活Twist1的表达,进而发挥其对EMT标记蛋白的转录调节。 Fan等[14]对193 例结直肠癌患者组织进行免疫组化检测发现,Twist1的异位表达可降低E-cadherin的表达,诱导EMT产生,从而促进癌细胞侵袭转移能力的增强,结合前面的实验,随着结直肠癌组织、高级别上皮内瘤变组织、低级别上皮内瘤变组织、癌旁组织,E-cadherin阳性率明显升高(P<0.05),而TGF-β1和Twist1阳性率明显降低(P<0.05),这表明Twist1、TGF-β1 的高表达以及E-cadherin的低表达在诱导结直肠癌细胞EMT过程中发挥一定作用,这一现象的发现可为我们后期探讨结直肠癌癌变的分子机制提供必要的理论基础。
本次实验结果我们发现结直肠癌E-cadherin、TGF-β1和Twist1表达在分化程度、Duke分期、浸润深度、淋巴结转移等方面均存在显著性差异(P<0.05),且结直肠癌E-cadherin与TGF-β1、Twist1均呈负相关性关系(P<0.05),TGF-β1与Twist1呈正相关性关系(P<0.05),这与国内外文献报道基本一致[10,13]。综上所述,E-cadherin、TGF-β1和Twist1三者与结直肠癌的发生发展等生物学行为密切相关,通过检测结直肠组织E-cadherin、TGF-β1和Twist1的表达有可能为判断结直肠癌的恶性程度及患者的预后提供线索并且有助于早期诊断结直肠癌和指导临床治疗。
猜你喜欢
中国现代医生(2022年19期)2022-08-25
中国临床医学影像杂志(2022年2期)2022-05-25
今日农业(2021年9期)2021-11-26
今日农业(2020年16期)2020-12-14
广东医科大学学报(2020年4期)2020-08-24
山东冶金(2019年2期)2019-05-11
数学杂志(2019年1期)2019-02-18
电影(2018年10期)2018-10-26
癌症进展(2016年9期)2016-08-22
中国卫生标准管理(2015年5期)2016-01-14
