
持续干旱对松叶百合保护酶活性的影响
2018-09-25 01:16程晓慧,褚昊天,陈少鹏,杨明明,程悠
吉林农业 2018年16期
关键词:干旱胁迫
程晓慧,褚昊天,陈少鹏,杨明明,程悠
摘要:选用3年以上的松叶百合(Lilium cernuum.)种球为实验材料,采用盆栽控水法进行模拟持续干旱处理,测定了不同干旱胁迫下松叶百合鳞茎保护酶活性。结果表明:干旱胁迫后,松叶百合外部鳞茎的SOD、CAT活性均随干旱时间的延长,呈现出先升高后降低的变化趋势,而POD活性则在干旱胁迫后期随时间的延长呈上升趋势。松叶百合内部鳞茎保护酶除了在干旱胁迫后期SOD活性有低于对照组的下降趋势外,其他4种保护酶活性均趋近于对照组。
关键词:松叶百合;干旱胁迫;保护酶活性
基金项目:吉林省高等学校大学生创新创业训练计划项目(项目编号: 吉农院合字〔2017〕第201711439006号)
中图分类号: S688.4 文献标识码: A DOI编号: 10.14025/j.cnki.jlny.2018.16.031
松叶百合(Lilium cernuum.)为百合科(Liliaceae)百合属(Lilium)多年生草本球根植物,又名垂花百合。松叶百合在1985年首次在云南被发现并命名[1],近些年随着多次对长白山珍稀植物资源的调查,在长白山余脉也发现了少量的松叶百合分布,且数量稀少。松叶百合植株低矮,极抗旱、抗寒、耐阴,花色为百合中少见的紫红色,是一种十分优良的园林绿化和鲜切花品种资源[2-3]。
然而,随着气候干旱加剧,城市园林用水量逐步增加,寻求耐旱的园林植物是节约城市环境用水的重要途径。植物细胞保护酶活性与植物抗逆性有密切关系,干旱胁迫条件下植物细胞内自由基产生和消除的平衡被打破,从而出现自由基积累,引发膜质过氧化作用,造成细胞膜系统破坏,直接影响植物正常生长,严重时导致死亡[4-6]。因此在长期进化过程中,植物为保护自身免受干旱胁迫伤害形成了相应的抗氧化保护酶系统[7]。本文从不同的干旱胁迫程度对松叶百合鳞茎保护酶活性的影响进行研究,分析松叶百合在干旱胁迫下体内保护酶活性的变化,以期为松叶百合在园林中的应用提供理论基础,同时利用人工栽培驯化,对松叶百合进行有效的保护。
1 材料与方法
1.1 供试材料
本试验在吉林农业科技学院教学实验基地进行,采用3年以上的松叶百合种球作为试验对象。
1.2 试验方法
本试验采用模拟盆栽法进行控水试验[8-10]。选取大小基本一致的3年以上的松叶百合种球为试材,用0.5%的多菌灵浸泡30min后,用蒸馏水清洗3遍,栽植于口径为21cm的花盆中,每盆一颗,共移栽150株。基质为园土与腐殖土4:1混合而成。盆栽苗生长稳定后,于5月中旬移入温室内,进行干旱胁迫处理。处理组一次浇透水后停止供水,对照组(CK)正常管理,浇水后每隔3d(0d、3d、6d、9d、12d、15d、18d、21d)取样,测定鳞茎SOD、POD、CAT活性,重复3次。
1.3 测定方法
酶活性采用南京建成SOD、POD、CAT快速检测试剂盒测定,方法参照说明书进行。
1.4 数据处理
数据均采用Excel 2010进行处理并绘制图表,用SPSS 19.0软件进行方差分析。
2 结果与分析
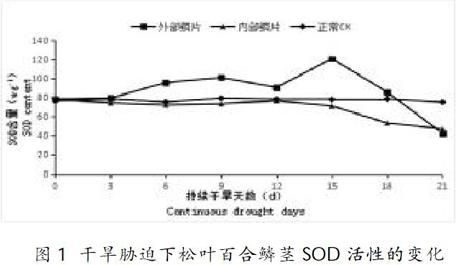
2.1 干旱处理对松叶百合SOD活性影响
超氧化物歧化酶(SOD)能有效清除外来胁迫产生的大量活性氧,其活性的变化能反映植物抗逆性的强弱[11]。从图1、表1可知,在干旱胁迫0~12d期间,松叶百合外部鳞片SOD酶活性先增高后降低;在12~15d期间,松叶百合外部鳞片SOD酶活性呈上升趋势,第15d时,松叶百合外部鳞片SOD酶活性达到最高,较CK增加了0.54倍,差异达极显著水平;在胁迫15~21d期间,松叶百合外部鳞片SOD酶活性呈下降趋势,在21d时,松叶百合外部鳞片SOD酶活性最低,但此时的SOD酶活性低于CK组,较CK组减少了0.43倍。在干旱胁迫持续过程中,松叶百合内部鳞片SOD酶活性一直低于CK组,但与CK相比较差异不显著;在15~21d期间时,松叶百合内部鳞片SOD酶活性呈逐渐下降趋势,在21d时,松叶百合内部鳞片SOD酶活性最低,较CK减少了0.37倍,差異达极显著水平。
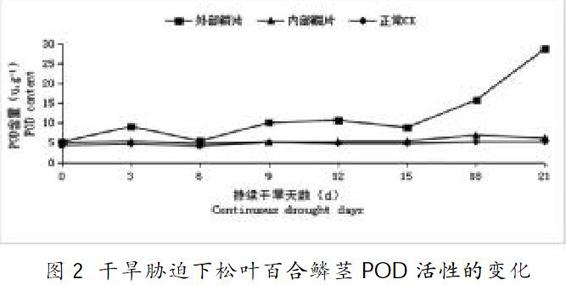
2.2 干旱处理对松叶百合POD活性的影响
过氧化物酶(POD)是植物体内清除活性氧的另一个重要酶类,可将干旱胁迫条件下POD活性变化幅度作为植物抗旱能力判断的生理指标[12]。从图2、表1可知,在干旱胁迫下,松叶百合POD活性整体呈现升高的趋势,在干旱胁迫0~6d期间,松叶百合外部鳞茎POD酶活性先增高后降低,6d时,松叶百合外部鳞茎POD酶活性与CK相比差异不显著;在干旱6~15d期间,松叶百合外部鳞茎POD酶活性呈小幅度增高后又降低,在15~21d期间,松叶百合外部鳞茎POD酶活性上升,在21d时达到最高,此时松叶百合外部鳞茎POD酶活性较CK组增加了4.51倍,差异达极显著。在干旱胁迫过程中,松叶百合内部鳞茎POD酶活性与CK相近,差异不显著,无明显活性变化趋势。
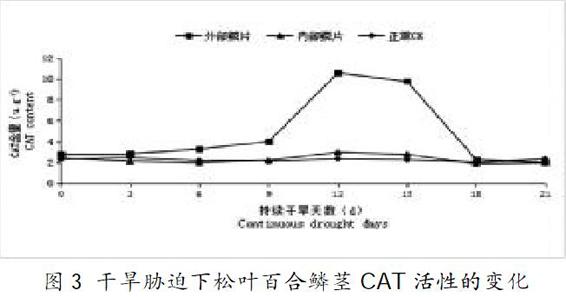
2.3 干旱处理对松叶百合CAT活性的影响
过氧化氢酶(CAT)催化细胞内过氧化氢分解,减少活性氧含量,使植株维持正常的生长发育[5]。从图3、表1可知,在干旱胁迫0~9d期间,松叶百合外部鳞茎CAT酶活性呈小幅度逐渐上升趋势;9~12d期间,上升幅度加大,在第12d时,活性达到最高,此时松叶百合外部鳞茎CAT酶活性与CK相比较增加了3.50倍[13],差异达极其显著水平;在12~18d期间,松叶百合外部鳞茎CAT酶活性呈大幅度下降,在18~21d期间,松叶百合外部鳞茎CAT酶活性下降,与CK相比差异不显著。在干旱胁迫过程中,松叶百合内部鳞茎CAT酶活性与CK相近,无明显活性变化趋势。
3 讨论与结论
干旱是限制植物生长和发育的主要逆境之一,干旱胁迫会造成植物体内产生大量的活性氧,有极强的氧化能力[9],积累的活性氧会导致膜质过氧化水平增高,破坏植物细胞膜完整性,导致植物组织伤害和衰老[15-16]。为了避免细胞膜遭受氧化伤害,植物需要建立一套抗氧化消除系统,有效清除自由基,其中包括SOD、POD、CAT这3种保护酶[17-18]。
通过试验对松叶百合进行21天干旱处理,干旱处理后鳞茎部分各保护酶活性的变化结果表明:SOD酶活性在0~15d期间,随着干旱时间的延长,SOD酶活性显著增加;在15~21d期间,SOD酶活性随着干旱时间的延长,酶活性大幅度降低,最后低于对照组。这与孙国荣[19]等在白桦树实生苗保护酶活性变化试验的研究结果一致。说明在一定干旱程度内,SOD酶活性具有有效清除活性氧和恢复适应的能力,后期可能是由于干旱超出了松叶百合鳞茎SOD酶的耐受限度,过多的过氧化物导致外部鳞茎受到过度损伤,合成SOD的能力下降,使SOD酶系统受到破坏。
松叶百合鳞茎POD酶活性随着干旱胁迫时间的延长呈逐渐上升的趋势,在干旱胁迫后期,POD酶活性达最高,这与邹原东[20]等人研究相符合。说明松叶百合外部鳞茎在严重干旱胁迫下仍能保持较高的POD酶活性,有效清除活性氧维持细胞膜的稳定性,避免膜脂过氧化伤害。从POD酶活性变化幅度可看出松叶百合具有较强的抗旱能力。
松叶百合鳞茎CAT酶活性均表现出先升高后降低的趋势,这与章崇玲[21]在干旱胁迫对菜苔叶片保护酶活性和膜质过氧化试验中结果一致。说明在干旱胁迫前中期,通过升高CAT酶活性来防止活性氧大量积累造成的氧胁迫毒害。但在胁迫后期,由于胁迫情况持续加深,3种酶的活性都急剧下降,此时松叶百合体内活性氧的形成和清除系统之间的平衡被打破,但都趋近于CK组。
研究表明,SOD、POD、CAT具有协同作用,可有效清除体内过多的自由基,提高了松叶百合的抗旱能力。松叶百合鳞茎外部4种酶变化明显,鳞茎内部变化趋势变化与CK相比较差异不显著,可能是因为随着干旱胁迫的加深,外部鳞茎优先受损严重,而内部鳞茎由于受到外部鳞茎的保护而受影响较小,到胁迫后期,整个清除自由基防御系统的防御能力减弱,推测可能是自由基过量产生,超出了防御系统的清除能力,使松叶百合进入休眠状态,地上部分也逐步枯萎。這说明松叶百合鳞茎在应对干旱胁迫时能通过牺牲外部鳞片从而保护内部鳞片的活性,以应对环境变化,是一种优良的耐旱植物。
参考文献
[1]彭隆金.百合属一新种——松叶百合[J].植物分类与资源学报,1985,7(03):317-318.
[2]郭太君,胡昕,葛新新,等.遮荫对细叶百合和松叶百合生长发育及光合色素含量的影响[J].东北农业大学学报,2013,
44(07):128-132.
[3]胡昕.干旱和遮荫对两种百合生长及生理特性的影响[D];吉林农业大学,2012.
[4]王莺璇,等.7种百合科园林地被植物的抗旱性研究[J].安徽农业科学,2012,40(26):12979-12983.
[5]季杨,张新全,彭燕,等.干旱胁迫对鸭茅根、叶保护酶活性、渗透物质含量及膜质过氧化作用的影响[J].草业学报,2014,
23(03):144-151.
[6]张旭颖,王玲玲,关,等.干旱胁迫对小叶黄杨幼苗膜脂过氧化及保护酶活性的影响[J].哈尔滨师范大学自然科学学报,2010,26(02):79-83.
[7]谢亚军,王兵,梁新华,等.干旱胁迫对甘草幼苗活性氧代谢及保护酶活性的影响[J].农业科学研究,2008,29(04):19-22.
[8]张鹏,孙明高,宋尚文,等.干旱胁迫对板栗幼苗保护酶活性的影响[J].山东农业大学学报(自然科学版),2010,41(01):6-10.
[9]宋礼毓,张兆斌,史作安,等.干旱对扁桃抗氧化酶活性变化的影响[J].落叶果树,2007(02).
[10]刘海燕,齐孝辉,韩雪,等.干旱胁迫及复水对四个白三叶品种叶片保护酶活性的影响[J].北方园艺,2010(05):104-
106.
[11]高亚梅,韩毅强,杜吉到,等.干旱胁迫对大豆酶活性的影响[J].黑龙江八一农垦大学学报,2007,19(04):13-16.
[12]李永华,翟敏,李颖旭,等.干旱胁迫下牡丹叶片光合作用与抗氧化酶活性变化[J].河南农业科学,2007,05:91-93.
[13]温玉龙,张文娟,邵长芬,等.高温和干旱胁迫对2 种景天植物膜脂过氧化及保护酶活性的影响[J].内蒙古农业大学学报(自然科学版),2014,35(3):37-40.
[14]郭雅玲,等.逆境下茶树叶绿体的理化特征研究进展[J].食品安全质量检测学报,2017,8(05):1610-1615.
[15]李忠喜,朱延林,张江涛,等.在干旱胁迫下饲料型刺槐无性系保护酶活性的差异[J].安徽农业科学,2007,35(10):2868-
2869.
[16]毕会涛,黄付强,邱林,等.干旱胁迫对灰枣保护酶活性及膜脂过氧化的影响[J].中国农学通报,2007,23(02):151-155.
[17]吴建华,张汝民,高岩.干旱胁迫对冷蒿保护酶活性及膜脂过氧化作用的影响[J].浙江林学院学报,2010,27(03):329-333.
[18]王霞,侯平,尹林克,等.土壤水分胁迫对柽柳体内膜保护酶及膜脂过氧化的影响[J].干旱区研究,2002,19(03):17-20.
[19]孙国荣,彭永臻,阎秀峰,等.干旱胁迫对白桦实生苗保护酶活性及脂质过氧化作用的影响[J].林业科学,2003,39(01):165-167.
[20]邹原东,韩振芹,高琼,等.干旱胁迫对葡萄保护酶活性的影响[J].北京农业职业学院学报,2016,30(01):31-34.
[21]章崇玲,曾国平.干旱胁迫对菜苔叶片保护酶活性和膜脂过氧化的影响[J].植物资源与环境学报,2000,9(04):23-26.
作者简介:程晓慧,在读本科生,研究方向:园林植物引种与栽培。
通讯作者:陈少鹏,博士,讲师,研究方向:园林植物引种与栽培。
猜你喜欢
湖北农业科学(2017年1期)2017-03-09
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
广西农学报(2016年3期)2017-02-06
安徽农学通报(2016年24期)2017-01-12
山东农业科学(2016年11期)2016-12-17
现代园艺(2016年2期)2016-03-15
江苏农业科学(2015年11期)2016-01-27
