
基于暂储理论的源头溪流营养盐滞留研究进展
2018-09-22 08:57黄钰铃骆辉煌毛战坡
中国水利水电科学研究院学报 2018年4期
黄钰铃,骆辉煌,马 巍,毛战坡
(中国水利水电科学研究院 水环境研究所,北京 100038)
1 研究背景
河流生态学研究中通常将平滩流量下河宽小于1.3 m、河长小于500 m的1-3级河流归为源头溪流(headwater stream)[1]。源头溪流是河流、湖泊、水库等水域系统的重要组成部分,其长度可达整个河流水系总长度的85%以上[2]。据调查,美国源头溪流总长度约占流域内河流总长度的70%~80%,汇集了约60%的水量及营养盐[3],在河流系统中占据举足轻重的地位。
我国河流众多,源头溪流数量庞大。农业生产、旅游开发、矿产开采、小水电建设等人类活动带来了污染物输入、河道破坏、河道减水脱水等问题;日益兴起的河道综合整治工程从防洪、景观等角度出发对河道进行了大量改造,忽视了上游河道对流域污染负荷输入的滞留与削减作用,这些都对下游的水环境情势和水生态系统造成不利影响。本文从暂储理论出发,系统梳理了源头溪流营养盐滞留研究进展,在此基础上辨析了目前研究中存在的不足,展望了未来的研究方向,以期为河流下游污染负荷削减、河流廊道生态恢复与重建提供支撑。
2 营养盐暂储及暂储区
2.1 暂储及暂储区定义流域内大量氮、磷汇集进入源头溪流中,随水流输向下游的过程中,一部分营养盐通过基质吸附、物理沉降、化学沉淀、生物吸收等作用滞留在源头河段[4-5],最终被分解利用或转化。这个滞留的过程称为暂态储存(transient storage,简称“暂储”),营养盐暂储的区域即为营养盐暂储区[6]。
营养盐暂储过程的研究始于1983年,Bencala等[7]从地下水-地表水相互作用的角度出发,提出了暂储模型(Transient Storage Model,TSM),用于定量描述山地溪流溶质的迁移规律。该模型针对溪流特点,定义了主流区和暂态存储区两个概念,模型由对流-扩散、暂态存储、侧向补给等模块组成。Runkel针对TSM模型的不足提出了主要用于描述氮、磷等营养盐迁移转化的模拟模型(One-Di⁃mensional Transport with Inflow and Storage,OTIS)[8]。营养盐暂储是溪流水域系统中影响溶质迁移的重要水动力学过程,不仅包括暂储区与主流区流动水体的交换作用,也包括暂储区内部溶质的滞留效应[9]。
2.2 暂储区类型暂储区主要包括两类:潜流暂储区(hyporheic transient storage zone,HTS)和表层暂储区(surface transient storage zone,STS)[10]。潜流区(hyporheic zone)是溪流河道中地表水与地下水双向迁移与混合、物质和能量交换与传递的区域[11-13]。潜流区孔隙较多,比表面积较大[14],可有效滞留营养盐[15]。表层暂储区主要指滨岸滞水区、基质裂隙、藻垫缝隙、回水区等[16-18],具有水陆交错、灌草丛生、生物膜黏附、水流回旋等特点,故能对营养盐产生不同程度的滞留作用。在对某山区小河中溶质输移的观测中,发现了5个表层暂储区,包括河床基质不规则产生的漩涡、深潭边的缓慢回旋水流、障碍物后的微型回旋水流、河岸裂缝涡流及粗基质裂隙涡流[7]。因此,识别源头溪流中不同类型的营养盐暂储区,是分析源头溪流中氮、磷等营养盐滞留过程的基础[19]。
2.3 影响暂储区的因素源头溪流中营养盐暂储区受许多因素影响,主要包括溪流水文条件、地形地貌特征以及生物因子,其暂储特性以暂储区断面面积As、营养盐滞留时间Ts、营养盐输移长度Ls、溶质交换系数αs、暂储时间中值Fmed等参数来表征[10]。
2.3.1 水文条件 营养盐暂储区受河流水文条件特别是河流流量的影响较大[20]。一般来说,表层暂储区溶质交换系数与溪流流量成正比[21];潜流暂储区交换系数受潜流区水力梯度和河床传导率影响,而水力梯度又受河床底坡、局部水流速和地下水循环等因素左右;营养盐滞留时间、暂储时间中值Fmed与溪流流量呈负相关[16]。暂储区断面面积与流量关系不明确[22-23]。对在1—6级河流中开展的246项示踪试验结果进行分析,发现暂储区面积与营养盐滞留时间随着河流等级增大而降低[24];另有研究表明暂储区面积与河流规模没有明显的联系[25]。
2.3.2 地形地貌特征 大量研究表明,河流规模、水流障碍物、深潭-浅滩、阶梯、碎石坝、弯道、支流等河床地形地貌控制着溪流暂储区的特征[4,26],从而影响营养盐的滞留能力[27]。基于对阿巴拉契亚山脉和蒙大拿州喀斯喀特山区河流的研究,估计得出营养盐暂储区与主流区的断面面积之比平均值为1.2,该比值在1级河流中最大,在5级河流中最小,在未受干扰的河段中达到0.6,在受干扰的河段中则为0.1[20]。在俄勒冈州威拉米特河上游的8级河流中,砾石床质的河床上有许多潜流暂储区,暂储区与主流区的断面面积之比平均为0.28[28]。在新罕布什尔州林区的13条源头河流中,营养盐暂储区与主流区的断面面积之比为0.16~0.71[29],略低于阿巴拉契亚地区河流中相应的值,与新罕布什尔州浅层河底生物带中的值大致相同。在高渗透性河床的河流中,暂储区与主流区的断面面积比值超过1[30]。在南极冰川融水补给的河流中,由于河床梯度较大、床质为多孔性冲积物,暂储区与主流区的断面面积之比超过1[31]。
河道内营养盐暂储区受石块和木头等障碍物的影响较大。在植被茂盛的农村河流和森林河流中,当植被和石块等障碍物被清除后,营养盐的暂储区面积分别下降了61%和43%[32];若重新投入障碍物,上述两类河流中营养盐的暂储区面积As值分别增加了3倍多和1倍多。
2.3.3 生物因子 溪流中潜流区具有较大的孔隙率及表面积,为生物提供了广阔的栖息生境[14]。据推测,美国蒙大拿州Flathead弗拉特黑德河潜流区生境范围是河道的2400倍[33]。潜流区中固着藻群落、基质上黏附的生物膜、大型植被等强烈影响暂储区的暂储特性[27,34]。在某些情况下,当河底潜流区较小、河水表层藻类和生物生产力较高时,营养盐暂储区与河床表面的联系较强。通过添加保守示踪剂氯化钠,发现在丝状藻和蓝藻生物量较高的实验河流中,暂储区较大;反之,在藻类生物量较低的河流中暂储区较小[17]。
3 暂储区对营养盐滞留的作用
3.1 营养盐输移
3.1.1 物理传输 溪流中溶质的输移过程可以对流-扩散方程描述如下[35]:

式中:C为溪流中的营养盐浓度;t为时间;v为溪流水流流速;D为扩散系数;x为输移距离。
上式用于描述保守粒子的对流-扩散过程。溪流中主要营养盐氮、磷为非保守物质,在描述其运动时,还应考虑迁入-迁出、生物吸收等导致的衰减作用。
3.1.2 营养盐螺旋 溪流中营养盐在主流区和暂储区中发生迁入-迁出,在暂储区中部分营养盐因吸附、吸收等过程发生滞留,随后通过解吸、释放等作用重新回到主流区并随水流输向下游,其输移过程可以下式表示[36]:

式中:C和Cs分别为溪流主流区和暂储区中的营养盐浓度;A和AS分别为溪流主流区和暂储区断面面积;α为营养盐在主流区和暂储区之间的交换系数;λc和λs为主流区和暂储区中生物对营养盐的吸收系数。其中,Cs和C的差值与营养盐扩散进出暂储区的速率成正比。
为了量化营养盐在溪流主流区和暂储区之间的输移特点,研究中引入了营养盐螺旋(nutrient spi⁃raling)的概念。河流中营养盐随水流输向下游的过程中,受河道内生物吸收和非生物滞留作用的影响,营养盐浓度变化较大,形成了营养盐螺旋[36-37](见图1所示)。营养盐螺旋长度S以吸收长度SW和周转长度SB两个参数来表达[38],吸收长度SW表示营养盐随水流传输的距离,周转长度SB表示营养盐通过生物吸收利用后分解释放回到水流中的距离。故有:
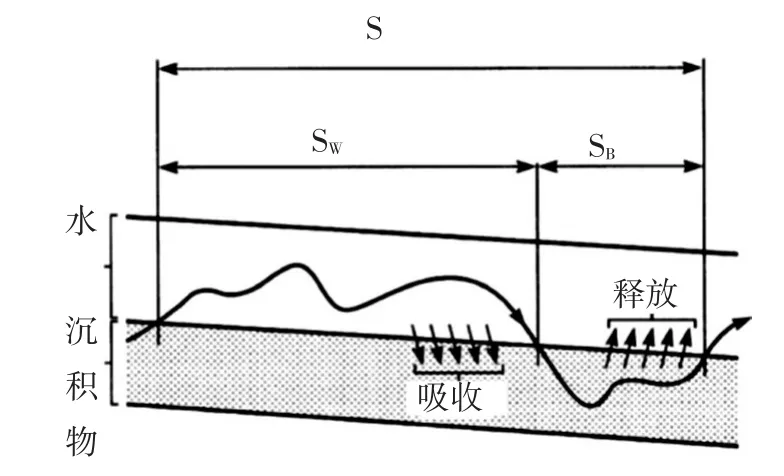
图1 溪流中营养盐螺旋

式中:S为螺旋长度;SW为吸收长度;SB为周转长度;k为营养盐综合衰减系数。
营养盐螺旋长度较短,表明溪流生态系统对营养盐需求旺盛,吸收利用率较高,营养盐发生滞留;反之,则表明河流生态系统代谢缓慢,对营养盐需求较低,营养盐易被输移至下游。营养盐吸收长度取决于河流流量和流速,而这些因素都与河流大小有关。可将SW标准化,转换为可衡量的溶质吸收速率vf,用来量化溶质通过生物或非生物过程从水柱到河床潜流暂储区转移的速率,其计算公式如下:

式中:vf为营养盐吸收速率;z为溪流水深。
将vf与溪流中营养盐浓度结合起来,可将单位时间内溪流潜流暂储区滞留的营养盐量表示为单位面积的吸收速率U,计算公式如下:

式中:U为单位面积营养盐吸收速率。
3.2 暂储区对营养盐滞留作用机理
3.2.1 理化反应 对潜流暂储区中营养盐滞留作用机理的研究较多。溪流中营养盐可能通过吸附—解吸、沉降—再悬浮等过程在水柱和潜流暂储区之间输移。吸附—解吸过程对于溪流中营养盐浓度起到了缓冲作用。当溪水中营养盐浓度过高时,潜流暂储区吸附营养盐使其沉降滞留下来;当溪水中营养盐浓度较低时,潜流暂储区的营养盐通过解吸作用被重新释放回到溪水中。在贝尔溪中开展的研究发现,夏秋季潜流暂储区是重要的氮储存库,沉积物吸附大量铵[39];数周乃至数月后,沉积物吸附的铵被解吸出来[3]。阿肯色州某河流接纳了污水处理厂出水后,潜流暂储区的沉积物吸附了大量磷[40]。加利福尼亚州某三级河流潜流暂储区中砾卵石床质沉积物可能是氨氮的临时储存库[41]。通过比较潜流暂储区沉积物和水陆界面河水中氨氮的含量,发现夏季基流中吸附的氨氮较高、冬季暴雨径流中吸附的氨氮较低。
除了吸附—解吸作用,在有氧条件下可溶性活性磷和溶解有机磷可能与金属氧化物和氢氧化物形成不溶性沉淀沉降至潜流暂储区,在厌氧条件下这些磷酸盐沉淀重新释放回水体中。
3.2.2 水文过程 溪流流量变化作用于暂储区从而可在不同时间尺度上对营养盐滞留产生影响。当流量较低时,磷营养盐吸附于细颗粒上,在颗粒表面形成生物膜,不易被水流挟带而滞留在暂储区中;在溪流流量较高的情况下,营养盐与沉积物和生物膜的接触时间缩短[3]。对北美11条河流中氨氮吸收长度SW进行对比时,发现营养盐吸收长度随河流流量和流速增加而延长[42]。在阿拉斯加库帕鲁克河流域中,随着河流规模增大,河流流速和水深增加,氨氮和颗粒态氮的吸收长度也随之增加[43]。在新西兰两条河流中也获得了类似的结果,氨氮、硝酸盐、磷酸盐的吸收长度随河流流速和水深发生显著的时空变化[44]。由于吸收长度SW强烈取决于河流流速和水深,研究中常以吸收速率vf来对吸收长度进行标准化转化以便比较。
研究发现,在河流中设置水流障碍物后,河水中氨和磷的吸收速率显著增加[32]。在新墨西哥州某河流中,营养盐吸收率和暂储区大小及河道流量成正比[45]。比较新罕布什尔州森林河流的13处上游源头时发现,夏季氨氮的吸收长度有35%与暂储区长度有关,年内平均来看有14%与暂储区长度有关,这可能是因为这些砾石河床上沉积物潜流区较多,营养盐的潜流暂储区较长[29]。
3.2.3 生物作用 暂储区中生物通过初级生产者同化吸收、异养生物异化转化、高等消费者摄食等方式促进营养盐滞留与去除。
(1)初级生产者同化吸收。亚利桑那州梧桐溪中氮的滞留取决于固着藻藻垫丰度,当固氮蓝藻占藻垫中固着藻的10%以上时,氮的滞留量较少[46]。北美11条源头溪流中氨氮主要通过自养生物和细菌、真菌的同化作用、沉积物的吸附作用以及硝化作用发生滞留并最终得以去除,其中沉积物吸附作用滞留去除70%~80%的氨氮,硝化作用滞留去除20%~30%的氨氮[3,42]。通过外源干扰杀灭水体中的微生物后,溪流中磷的吸收长度增加了一倍多[47]。在阿巴拉契亚山脉地区源头河中,当移走河水中的枯枝落叶和倒木后,营养盐吸收长度大幅增加[48]。比较两条河流发现,生物群落呼吸作用较强的溪流中磷的利用率高[49]。
溪流中水文条件变化后,暂储区初级生产者的丰度等也随之改变,影响了营养盐滞留。南极河流流量出现极端值时,丝状蓝藻优势种群发生变化,营养盐滞留随之而变[50]。地中海森林河流中,磷的滞留主要取决于藻类,而氨氮的吸收速率与藻类活动的联系很小,其主要是受微生物异养过程影响[51]。在新西兰两条草原河流中,营养盐的吸收速率在春季最高,在秋季和早冬最低,这主要由藻类丰度变化所致[44]。
着生有大型水生植物的表层暂储区对营养盐的滞留作用很强。在对新罕布什尔州某河流的研究中发现,河水流经苔藓植物河床时,大量磷被滞留去除[52]。在田纳西州沃克河支流中,投入示踪剂15N,6周后发现苔藓植物吸收的氨氮占总滞留氮的41%[53]。同理,德国施普雷河中大型植物床延缓了水流,延长了水力停留时间,促进了有机物沉积并显著增加了磷的滞留量[54]。
(2)异养生物异化转化。营养盐通过异养生物的作用发生形态转化,最终从溪流中去除。密歇根州鹰溪和波多黎各比斯利山区溪流中硝化菌将约50%以上的氨氮滞留、转化、去除,但在田纳西州沃克河支流中仅滞留、转化、去除了20%左右的氨氮[54]。农业地区河流中,反硝化菌脱氮作用较强,促进了氮的滞留与去除[55-56]。在低速流和浅水区的潜流暂储区中,反硝化菌脱氮速率很高,可去除大量硝酸盐,如在田纳西州源头河流中,反硝化脱氮占总滞留、去除氮的16%[53]。在美国东北部地区河流内37%~76%的氮通过反硝化菌脱氮作用去除,且一半发生在1~4级河流中[35]。科罗拉多西南部东河中潜流暂储区提高了反硝化菌脱氮速率,由此导致氮在源头河段滞留后被分解释放,降低了对下游的输出[57]。
(3)消费者摄食。捕食者通过影响底栖生物和有机物、捕食藻类来直接影响营养盐滞留;腐食者将粗颗粒有机物质破碎后摄食利用,促进营养盐滞留[58]。当藻类和生物膜被大量摄食后,营养盐的吸收率降低,营养盐螺旋长度加长;草食动物适度的取食会刺激初级生产者和微生物,因此会产生相反的效果。在实验室人工河流中,蜗牛被移除后,对藻类的摄食压力降低,藻垫由此蓬勃生长并逐渐密实,形成了营养盐暂储区,溪流中营养盐螺旋距离变短,营养盐滞留并被吸收利用[17]。
4 当前研究存在的问题
(1)源头溪流中营养盐滞留与暂储区之间的联系尚不明确[10]。从当前资料来看,不同种类营养盐受暂储区的滞留作用差异较大,同种营养盐在时空条件不同的暂储区中的滞留过程也有很大差别。研究发现,林地溪流与草地溪流中活性磷营养盐与暂储区正相关,但两类溪流中氨氮的吸收速率均较低且沿程变动较大,表明对氨氮的滞留作用很弱;与林地溪流相比,草地溪流暂储区更大,对活性磷营养盐的吸收长度更短、吸收速率更高[59]。某些情况下,有些溪流中暂储区对营养盐的滞留作用很小。在冰川融雪溪流中,潜流暂储区对正磷酸盐的滞留作用微弱[50]。美国梧桐溪中营养盐吸收长度和暂储区之间没有相关性[46]。在美国十条未受干扰的自然河流中氨的吸收速率和暂储区之间也没有相关性[42]。比较发现新西兰三条河流中可溶性活性磷和硝酸盐的吸收长度也与暂储区没有明显的相关关系[60]。在少数研究中甚至发现随着暂储能力的增加,磷的吸收速率不升反降的现象[61]。因此,在具体研究过程中,因流域水文条件、地形地貌及生物化学等因子的不同,源头溪流中营养盐滞留与暂储区之间的联系尚需进一步明确。
(2)在干扰驱动下暂储区变化对营养盐的滞留作用方面的研究较少。当流域自然环境变化如干旱、洪水、滑坡等事件发生时,源头溪流水文条件和地形地貌发生极大改变,随之对溪流中各类暂储区的水文水动力条件及生物地球化学特性产生极大冲击。此外,人类活动如矿产开采、旅游开发、小水电建设、农业活动、河道整治等将强烈干扰源头溪流中各类暂储区。自然和人为外源干扰作用下源头溪流中暂储区改变驱动其暂储特性发生变化,由此,溪流中营养盐滞留对暂储区特性变化的响应过程也随之发生改变[27]。当前研究仍多集中在暂储区特性改变后营养盐滞留过程的变化等“末端”,对暂储区特性如何受到影响、哪些影响因素可调控等“前端”的研究还较少。
(3)对通过人工调控促进营养盐滞留的水环境保护意义认识不足。目前,国内学者研究了水库、湖泊等对河流营养盐的滞留[62-67]、生物滞留系统对流域面源污染负荷的截留与控制[68-69]、农田溪流系统中污染物的滞留等[70-73]。个别学者开始关注源头溪流中的营养盐滞留问题[74-76],并在巢湖流域开展了一些工作[77-79],取得了初步成果。上述研究为评估河流筑坝的生态效应、预测湖库富营养化趋势、削减面源污染负荷等提供了大量基础数据。但研究中缺乏对数量庞大的源头溪流受纳的陆域营养物质滞留作用的重视。通过主动开展适宜的人类活动,改变源头溪流中的暂储区类型及分布,从而驱动营养盐的滞留,可实现两方面的目标:一是促进营养盐循环、维持源头河段初级生产力及生态系统动态平衡,二是削减河段中短期内富集的营养盐、特别是营养盐背景值较高的矿区河流及重污染的农业河流下游的入库污染物负荷。
5 结论与展望
综上所述,当前关于源头溪流营养盐滞留的研究已从暂储理论出发,通过实地观察与数学模型得出了一系列成果。研究表明水文、地形地貌及生物因子通过单因素或综合作用于暂储区,左右其暂储特性,并由此驱动暂储区内各类营养盐的理化反应、水文过程及生物吸收与利用。当前来看,对暂储区在外源干扰作用下的系列变化及暂储区内营养盐滞留的响应过程等方面的研究尚有不足;对调控人工干扰,调节暂储区分布,驱动营养盐滞留,促进营养盐就地消纳,维持源头河段生态平衡,削减下游污染负荷的系统性全盘考虑仍很匮乏。
对源头溪流中营养盐滞留特征和滞留机制的研究已成为环境科学、环境生态学、环境水文地质学等领域的研究热点和前沿课题。未来有必要从以下方面开展一些工作:(1)选择典型流域开展源头溪流营养盐滞留研究,以营养盐暂储理论为基础,探索不同营养盐滞留与暂储区之间的内在联系,丰富营养盐暂储理论;(2)结合当前自然变化或人类活动对流域干扰作用较强的现实条件,通过多种手段揭示暂储区在受到干扰、发生变化后影响溪流中营养盐滞留的机理;(3)选择流域营养盐高背景或人类活动强干扰的典型河流,在源头溪流中开展多情景调控,细致调查暂储区的变化,分析营养盐滞留过程,评估源头溪流下游控制断面营养盐负荷削减程度,为下游流域污染负荷削减、整个河流廊道的生态恢复提供理论支撑。
猜你喜欢
环境工程技术学报(2022年3期)2022-06-05
风流一代·经典文摘(2021年10期)2021-10-25
意林(2021年13期)2021-07-29
科学导报·科学工程与电力(2019年24期)2019-09-10
艺术评鉴(2019年9期)2019-06-17
水能经济(2018年7期)2018-07-12
小溪流(画刊)(2017年12期)2018-01-10
理论观察(2016年11期)2017-04-17
中国海洋大学学报(自然科学版)(2015年5期)2015-03-20
中国水利(2015年2期)2015-02-28
