
同域分布大熊猫和水鹿生境利用分异特征
2018-09-19 08:26白文科黄金燕张晋东夏珊珊周材权
生态学报 2018年15期
王 盼,白文科,2,黄金燕,张晋东,*,刘 巅,夏珊珊,饶 佳,周材权,2
1 西华师范大学珍稀动植物研究所 西南野生动植物资源保护教育部重点实验室,南充 637002 2 西华师范大学生态研究院,南充 637002 3 中国大熊猫保护研究中心,卧龙 623006
大熊猫是世界生物多样性保护的旗舰物种,也是中国特有珍稀孑遗动物。为保护这一珍稀濒危物种我国政府自20世纪60年代开始建立自然保护区,迄今为止全国已建立以大熊猫为主要保护对象的自然保护区67个,保护大熊猫栖息地258万hm2[1]。在开展大熊猫及其栖息地保护的同时,由于其产生的“伞护效应”也保护了与其同域分布珍稀濒危动物及其栖息地,其中有超过96%的大熊猫栖息地与其他特有两栖类、鸟类和哺乳动物的分布区相重叠[2]。但是,保护区内过度采伐迹地的森林植被目前并未完全恢复,缺乏完全的水文功能,生物多样性丰富度差[3],再加上放牧等人类活动的影响,栖息地丧失和破碎化仍旧是对大熊猫及其同域分布物种生存的重要威胁[4- 6]。
关于大熊猫(Ailuropodamelanoleuca)生境的研究主要涉及特定区域、特定季节、不同森林起源上的大熊猫生境选择和利用,研究对象大多是针对大熊猫单一物种的研究[7- 13]。但是在大熊猫栖息地保护和生境恢复的进程中,这种将单个物种作为保护对象,极具针对性的保护计划的效果已经受到质疑,需综合考虑多个物种对生境的需求,才能制定出更科学的保护计划[14]。国内已经有学者关注到了这一问题并开展了大熊猫同域分布物种的调查,主要涉及大熊猫伴生动物种类调查、种群分布、动态变化等较为基础的研究[15- 18],但是仅有少数研究分析了大熊猫与其同域分布物种在生境选择与利用中的异同[2,19-22]。
卧龙自然保护区是我国最早成立的以保护大熊猫为主的国家级自然保护区,保护区内有着丰富的动物资源,研究表明与大熊猫同域分布的野生动物有小熊猫(Ailurusfulgens)、川金丝猴(Rhinopithecusroxellanae)、扭角羚(Budorcastaxicolor)、水鹿(Rusaunicolor)、斑羚(Naemorhedusgoral)等多种珍稀兽类,其中水鹿是在大熊猫核心栖息地内分布最多、野外遇见率最高的有蹄类之一[23]。水鹿和大熊猫都是主要的林栖动物,有研究发现卧龙自然保护区内水鹿主要在森林和竹林活动,并且水鹿会采食冷箭竹的竹叶[24],这表明水鹿和大熊猫不仅在生态位上有着重叠现象,二者还存在着竞食关系,但国内目前并未有关于大熊猫与水鹿的共存机制研究,本研究首次将大熊猫和水鹿的空间利用和生境选择特征进行比较,以探究同域分布动物生境利用分异特征。对同域分布的野生动物开展生境和空间利用研究是生物多样性保护的理论基础,有利于更深入的了解物种对空间和资源的竞争与共存机制。
1 研究方法
1.1 研究区域
四川卧龙国家级自然保护区(102°52′—103°24′E、30°45′—31°25′N)始建于1963年,是我国建立最早的以保护大熊猫及森林生态系统为主的综合性自然保护区之一,保护区东西长52 km,南北宽62 km,总面积达2000 km2。保护区处于成都平原向青藏高原的过渡地带,海拔变化显著,山高谷深,生态系统类型多样。主要有皮条河、正河、西河和中河,河流两侧发育有许多支流,形成树枝状水系[25]。
保护区内的核桃坪、牛头山和五一棚区域毗邻皮条河,东南面由老鸦山河谷包围,海拔1840—3200 m,森林植被类型多样,生长有拐棍竹(Fargesiarobusta)、冷箭竹(Bashaniafangiana)和短锥玉山竹(Yushaniabrevipaniculata),是大熊猫活动核心区[26]。长期野外工作发现这些区域分布有大量的水鹿活动痕迹,故选为研究区域(图1)。

图1 卧龙自然保护区野外样方设置点Fig.1 Location of field sample in Wolong Natural Reserve
1.2 样方设置
本研究将研究区域内不同海拔上的兽径作为研究样线,在每条样线上每隔200 m做一个植被样方。每个乔木样方设置为20 m×20 m,记录坡度、坡向、动物痕迹(如食迹、粪便、实体等)等;在乔木样方内,以乔木样方中心点沿坡向的水平垂直线与乔木样方边界为界设置4个10 m×10 m的灌木样方,记录灌木盖度、平均高度、密度等;此外,在乔木样方内成“品”字形设置3个竹子样方,竹子样方之间直线距离至少大于6 m,记录竹子盖度、密度、高度、基径等(表1)。
1.3 分析方法
1.3.1 空间利用
为量化水鹿和大熊猫的空间利用竞争特征,本研究采用位点重叠计算公式计算两种动物的空间重叠关系。空间重叠系数计算公式为[27]:
OI=Oij/(Oi×Oj)1/2
式中,OI表示重叠度系数;Oij表示水鹿和大熊猫共同出现的样方数;Oi、Oj表示:有水鹿(大熊猫)出现的样方数。
1.3.2 生境选择
本研究随机选取了不同森林起源的生境做植被样方调查,以计算大熊猫和水鹿对原始林和次生林生境的利用强度。在每个样方中选择并记录了27种生境因子,用以分析水鹿和大熊猫对不同生态因子的选择差异,先用单一样本Kolmogorov-Smirnov 检验对数据进行正态分布判定,对满足正态分布的数据采用独立样本T检验进行运算,非正态分布的数据采用Mann-Whitney U检验进行运算。检验皆使用SPSS 22.0软件进行,显著水平为0.05。

表1 野外样方生境因子基本信息
2 结果与分析
2.1 空间利用

表2 同域分布大熊猫和水鹿生境空间重叠系数
在研究区域(核桃坪、牛头山、五一棚)共调查了400个样方。其中有大熊猫痕迹的样方263个,有水鹿痕迹的样方222个,同一样方中大熊猫和水鹿痕迹都有的样方有141个。利用位点重叠公式计算得出在大熊猫核心栖息地中大熊猫和水鹿的位点重叠系数为58.35%,在原始林生境中的重叠系数高达66.58%,如表2。这一结果表明在大熊猫核心栖息地中水鹿和大熊猫的生境重叠范围较广,二者在原始林生境中的空间竞争较大。
2.2 生境选择
根据大熊猫和水鹿样方数在森林起源上的不同,分析二者对原始林和次生林的利用强度。如图2,结果表明两种动物都在原始林的出现率较高,差异在于大熊猫对原始林的利用率高达82.89%,而水鹿对原始林的利用率为67.12%,表明大熊猫对原始林的依赖性更强。
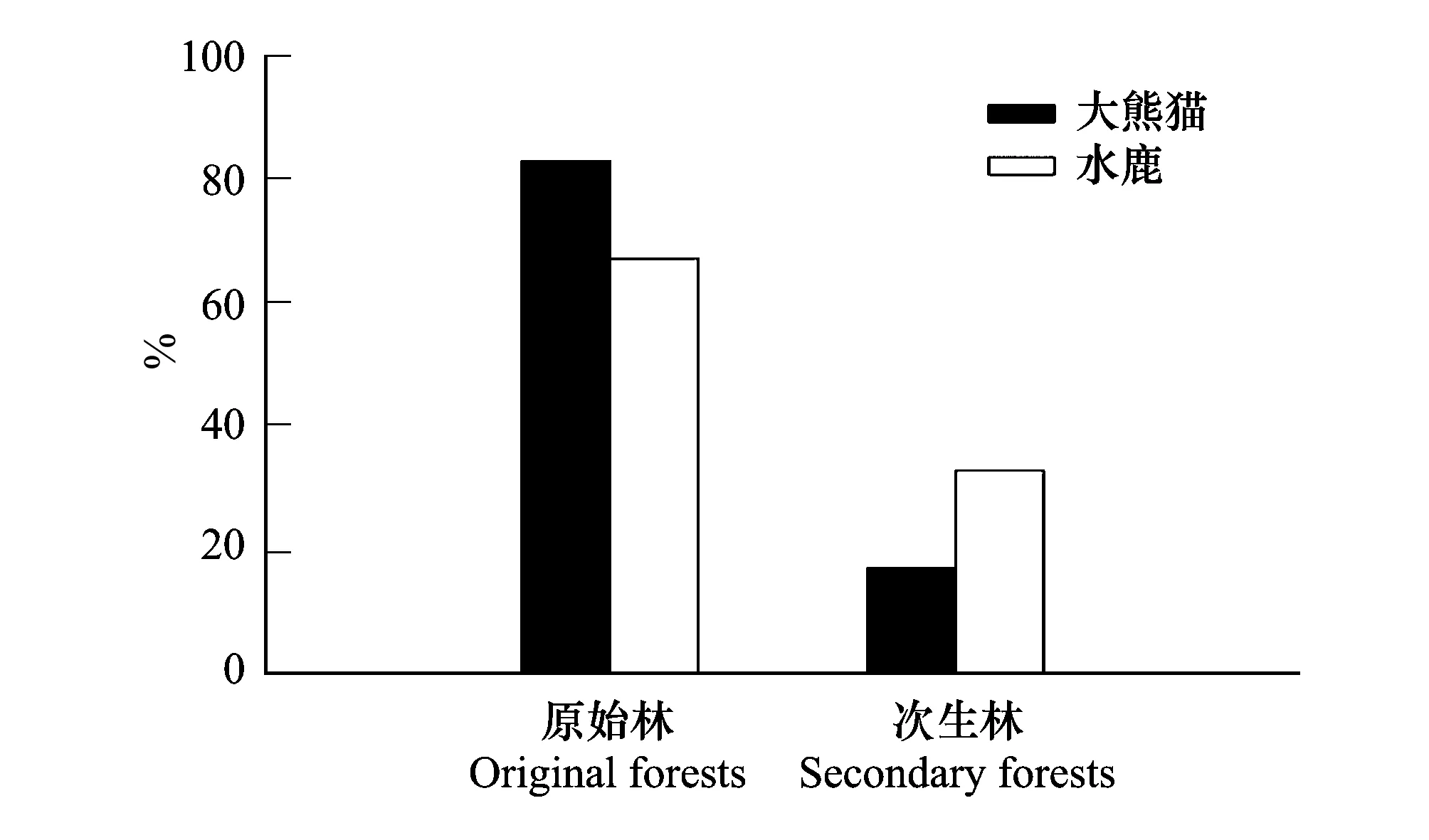
图2 同域分布大熊猫和水鹿原始林和次生林的利用率比较 Fig.2 Utilization rate Comparison of original and secondary forests of sympatric Giant Panda and Sambar
在调查过程中仅有大熊猫痕迹的样方有122个,仅有水鹿痕迹的样方有81个。对5个物理因子进行差异性分析,结果如下表(表3),坡位(P<0.01)、离小路距离(P<0.05)、离水源距离(P<0.05)有显著性差异。大熊猫出现率较高的区域位于中坡位和上坡位、离小路距离小于300 m、离水源距离小于500 m的地方;而水鹿出现率较高的区域位于中坡位和下坡位活动、离小路200—300 m、离水源距离小于300 m的地方。
对22个生物因子进行差异性分析,结果显示(表4):大熊猫和水鹿对乔木层因子中的乔木密度(P<0.05),灌木层因子中的灌木盖度(P<0.01)和灌木密度(P<0.01),竹林因子中的竹林盖度(P<0.05)、幼竹密度(P<0.01)、幼竹基径(P<0.05)幼竹高度(P<0.05)、成竹高度(P<0.01)、死竹密度(P<0.01)的选择有显著性差异。水鹿在乔木密度较高、灌木盖度为50%—90%、密度适中的区域出现率较高;大熊猫在灌木盖度较高(>75%)但是密度较低、竹林盖度和幼竹密度高、幼竹基径小、死竹密度低的生境出现率较高,在幼竹高度和成竹高度变量的选择中大熊猫呈现随机选择的趋势;而水鹿对竹林层因子的选择无显著性规律。
3 讨论
从空间利用上分析,大熊猫和水鹿的空间重叠系数高达58.35%,在原始林中的空间重叠率为66.58%,说明二者在空间分布格局上具有重叠现象,在原始林中空间资源的限制会导致二者的生态竞争加强。对大熊猫而言,原始林生境中高大的乔木可以为大熊猫提供育幼、觅食、躲避天敌、远离干扰等条件,有利于大熊猫的生存繁衍[28]。对水鹿而言,原始林和次生林处于不同的植被演替阶段,次生林中的灌木和草本生长旺盛密度较大[7],会影响水鹿在林中的活动,所以水鹿在原始林中的出现率较高。

表3 大熊猫和水鹿生境利用物理因子检测结果

表4 大熊猫和水鹿生境利用生物因子检测结果
*表示P<0.05,差异性显著;**表示P<0.01,差异极显著
从森林起源上分析,大熊猫对原始林的利用率为82.89%;对次生林的利用为17.11%,而水鹿对原始林的利用率为67.12%;对次生林的利用为32.88%,说明在对原始林和次生林的利用中水鹿与大熊猫具有相似的利用偏好,但是相对于大熊猫而言,水鹿对次生林的利用率更高。已有研究表明卧龙自然保护区的次生林生境经过几十年的保护已经得到了较大程度的恢复,具备大熊猫生境选择的基本条件[13]。但是本研究结果显示,即使在已经具备大熊猫生存条件的次生林生境,在实际应用中大熊猫对次生林生境的利用率相对于原始林的利用率还是很低,大熊猫依然更偏好在原始林中活动。
微生境的分离是同域分布物种共存的主要原因[29]。野生动物选择栖息地时会考虑到其他物种的出现和介入[30],大熊猫对生境因子的选择具有不同程度的可塑性和灵活性[31],为避免与同域分布动物在空间上的剧烈竞争,大熊猫在物理因子的选择上与水鹿产生了分异,二者分别利用不同坡位和离小路不同距离的生境使生态竞争降低。有研究表明水鹿几乎常年在水流附近活动[32],大熊猫也喜欢离水源较近的生境[20,33],但是本研究发现水鹿对水源的依赖性更强,水鹿几乎常年离不开水[32],而大熊猫在非采食竹笋的季节才会通过大量饮水来弥补体内水分的丧失[34]。
大熊猫和水鹿在生物因子的选择中,有乔木密度、灌木盖度、灌木密度、竹林盖度、幼竹密度、幼竹基径、幼竹高度、成竹高度和死竹密度等9种生态因子有显著性差异。动物的耐受性定律表明动物对每种生境因子的选择有一定的范围以满足其生存需求[35],竹林作为大熊猫觅食的主要活动区域,大熊猫对其各因子的选择范围更为明显,因为较高的竹林盖度可为大熊猫提供良好的隐蔽条件,幼竹的生长状况与大熊猫的食物直接相关;而灌丛作为水鹿觅食的主要活动区域,水鹿在灌木盖度适中(50%—90%)的生境出现率较高,且水鹿对灌木密度的适应选择范围更大。但是,这种生境差异性选择并非单一因子影响,例如水源地周围的坡度较为平缓,灌木种类多样性较为丰富,而水鹿主要以各类灌木嫩枝叶为食[32,36],灌木越多的区域水鹿的食物越充足,聚群生活的水鹿需要食物丰富的生境以满足群体对能量的需求。
野生动物理想的适宜生境需要包含各种功能斑块,能够满足其生存和繁殖所需要的一系列条件[37]。对同域分布的大熊猫和水鹿而言,二者在空间利用上有较高的重叠现象,但是对微生境的选择和利用方式有显著性差异。两种动物选择了更能满足各自生理和生态需求的生境,以满足自身对能量的需求,并减少在空间分布上有较高冲突时所产生的生态竞争,说明二者对生境的利用存在差异性,具有独特的共存机制。野生动物保护最终的目标就是维持生物多样性[38],卧龙自然保护区具有长达几十年的保护历史,在“退耕还林”和“天然林保护工程”等政策的推动下,其生境质量明显提高,动植物数量也有明显增长的趋势。研究同域分布物种在面临资源和空间上竞争压力时的生境利用关系,是开展物种多样性保护研究的重要步骤。所以,有必要继续开展大熊猫同域分布物种的生境利用关系和共存机制研究,这对保护区制定重要珍稀濒危物种的保护政策具有重要意义。
猜你喜欢
林业勘查设计(2022年1期)2022-02-15
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
落叶果树(2021年6期)2021-02-12
黑龙江工程学院学报(2020年5期)2020-10-21
林业调查规划(2020年3期)2020-06-03
水土保持研究(2019年6期)2019-10-19
体育科技(2016年2期)2016-02-28
海南热带海洋学院学报(2015年5期)2015-03-14
西华师范大学学报(自然科学版)(2015年3期)2015-02-27
