
景观水体周丛生纤毛虫群落结构及其与环境因子的关系
2018-09-19 08:28:30吴晓敏郝瑞娟王丽卿潘宏博
生态学报 2018年15期
吴晓敏,郝瑞娟,王丽卿,潘宏博,*
1 上海海洋大学水产科学国家级实验教学示范中心, 上海 201306 2 上海海洋大学农业部鱼类营养与环境生态研究中心, 上海 201306 3 上海海洋大学水产种质资源发掘与利用教育部重点实验室, 上海 201306
与高等真核生物相比,原生动物为单细胞动物,它分布广泛,能更直接与它所生存的环境接触,对环境变化如水体污染的反应更敏感[1]。因此,在水质监测中,原生动物可作为水体污染评价理想的环境指示生物[2- 6]。
周丛生纤毛虫是水域生态系统的重要组成部分,其在微食物网和生态系统的物质循环以及能量流动中起着非常重要的作用[7- 10]。纤毛虫生长速度快,可短时间在人造基质(如载玻片)上群集并获得较高的密度和丰度,成为周丛生群落的主要类群[11]。自Cairns等[12]首次用聚氨酯泡沫(PFU)法采集微型生物群落,周丛生纤毛虫已被广泛地用于生态调查的水质评估和监测中[11, 13- 14]。
目前用于生物监测的周丛生纤毛虫采样法主要是PFU法和载玻片法两种。沈蕴芬等[15- 16]对原生动物群落进行详尽的研究,并对PFU法进行改进和完善,将其发展成国标,从而用于测试水体的污染。近十几年来,PFU法广泛应用于淡水河流、湖泊和海洋的水质监测[5, 17- 20],载玻片法多应用于海洋监测[4, 21- 22]。由于景观水体面积小、水浅,PFU上的小孔极易被搅起的底质或大颗粒悬浮物堵塞而造成群落结构失真,无法正确监测水质,载玻片法则可以避免这个问题。基于载玻片法的生物监测在淡水尤其是景观水体研究极少,其上生长的纤毛虫等原生动物群落能否较好反映景观水体水质还有待确定。本研究于2015年1—12月在上海市东南角的一处浅型景观水体中利用载玻片法采集周丛生纤毛虫,并分析其群落结构与环境因子的关系,以期为周丛生原生动物的生态学研究提供第一手参考资料;同时,为评估景观水体的负荷力和能量流动规律之间的关系、建立景观水体中各个相关功能类群的动力学参数以及景观水体的水质监测等方面提供重要的资料。
1 材料与方法
1.1 采样时间和地点
2015年1—12月逐月采集上海市东南角一处人工景观水体(地理坐标为30°53′11.01′′N,121°53′26.85′′E)的周丛生纤毛虫。该水体位于上海南汇东滩湿地,横穿上海海洋大学校园,其一端有闸口,外接芦潮引河直入东海。总水体面积约为0.061 km2,平均水深约为3.20 m,海拔约为4 m[23]。
1.2 样品采集与处理
本研究利用载玻片(2.6 cm×7.6 cm)作为人工附着基进行景观水体周丛生纤毛虫的采集,使用前将载玻片浸泡在酒精中24 h后进行燃烧去除其表面脂结构。每个月放置8个载玻片框(112张载玻片)于湖中,采集的载玻片框(每框内置14片载玻片,两两背对着放置于框内的一个玻片槽中)悬挂于水深1 m处(载玻片框装置参考Xu等[4]),待周丛生纤毛虫群落达到成熟(春、夏、秋季5—8 d,冬季12—14 d)后将载玻片框取出,并放于原位水中带回实验室。
1.3 种类鉴定与定量研究
在体视显微镜下对一个载玻片框中的14张载玻片进行全片观察(65×)以获取种类数;随机选取2张载玻片,每张随机选取10个视野进行物种密度的统计[4],所有的计数均在12 h内完成[4]。种类鉴定主要参考文献[16, 24-29]。
1.4 水质参数的测定
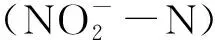
1.5 数据统计和分析
1.5.1 优势度的计算

1.5.2 多样性指数的计算
选取Shannon-Wiener 多样性指数(H′)[32],Margalef 丰富度指数(D)[33]和Pielou 均匀度指数(J′)[34]来分析周丛生纤毛虫群落结构的多样性。
Shannon-Wiener 多样性指数(H′):
Margalef 丰富度指数(D):
D=(S-1)/lnN
Pielou 均匀度指数(J′):
J′ =H′/lnS
式中,Pi=Ni/N,Ni为第i物种的个体数,N表示样品中所有种类的总个体数,S为样品中物种的总种类数。
1.5.3 分析软件
本研究数据的处理和分析采用SPSS 18.0软件进行Pearson相关性分析;折线和柱状图均采用OriginPro 2016绘制;采用Canoco for Windows 4.5软件对周丛生纤毛虫优势种的密度进行ln(X+1)转化,并将其与环境因子进行冗余分析(Redundancy analysis,RDA);采用PRIMER 6软件对周丛生纤毛虫密度进行平方根转化、对环境变量进行Log(X+1)转化并进行聚类分析(CLUSTER)以及生物-环境变量的相关性分析(BIOENV)。
2 结果与分析
2.1 水体环境因子的周年变化
本研究中,水体高锰酸盐指数(CODMn)的年均值为(6.82±1.04) mg/L,变化范围为5.22—8.58 mg/L,5月最低,8月最高;叶绿素a(Chl-a)浓度的年均值为(0.02±0.02) mg/L,变化范围为0.01—0.07 mg/L,8月浓度最高。
透明度(SD)年均值为(98.50±8.98) cm,其变化范围为80—110 cm,1月透明度最低,最高值出现在8、10和12月;水温(T)的平均值为(19.14±7.66)℃,变化范围为7.60—29.60℃,全年水温整体呈先升高后降低的趋势,最低水温出现在2月,最高水温出现在7月;溶解氧(DO)的年均值为(6.64±3.45) mg/L,变动范围为2.44—11.01 mg/L,6月最低,4月最高;电导率(Spc)的年均值为(971.09±182.57) μs/cm,变化范围为750.00—1282.00 μs/cm,9月最低,6月最高;年均盐度(Sal)为(0.56±0.07)‰,变化范围为0.44—0.64‰,10月最低,5月最高;酸碱度(pH)年平均值为(7.75±0.31),在7.15—8.35之间变动,11月最低,2月最高,表明本研究水体的水质酸碱度呈中性偏碱性(表1)。
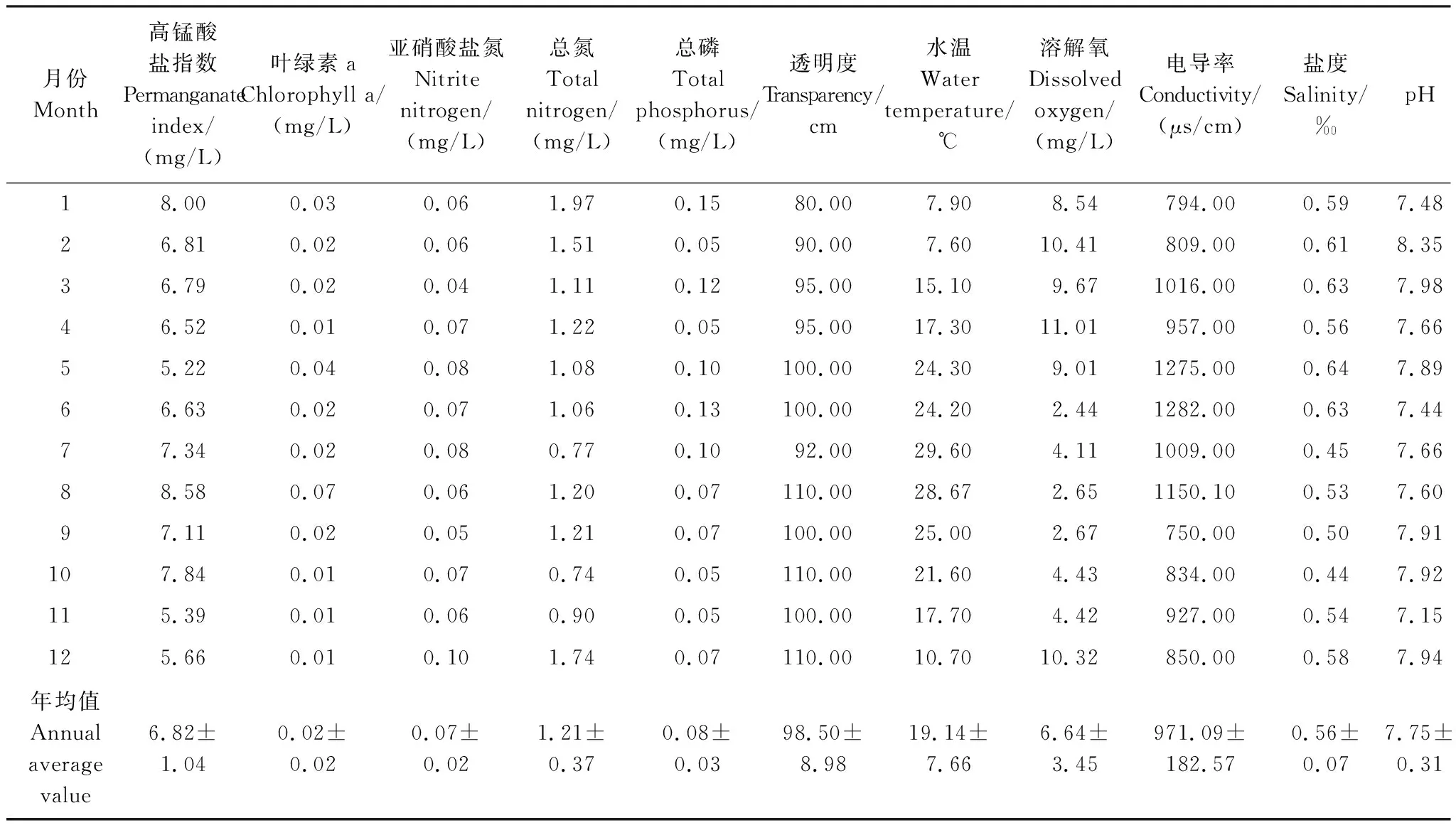
表1 每月理化因子数据
2.2 周丛生纤毛虫群落结构的动态变化
2.2.1 密度及种类数的周年变化
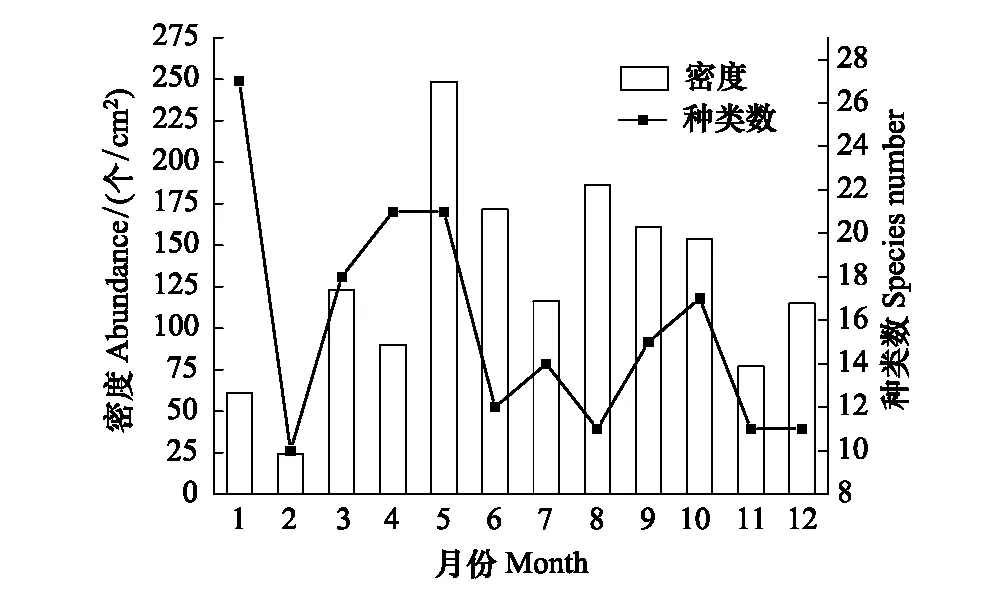
图1 周丛生纤毛虫的种类数及密度的周年变化 Fig.1 Annual variations of species number and abundance of periphytic ciliates
周丛生纤毛虫的年平均密度为127.29 个/cm2,年密度变化在24.27—248.57 个/cm2之间,2月密度最低,密度最高峰出现在5月,在8月出现次高峰。在全年的研究中,共鉴定周丛生纤毛虫51种,隶属于12目。种类数为春季>冬季>夏季>秋季。1月检出的周丛生纤毛虫种类最多,2月种类数最少,并且到5月一直处于增加的趋势,变化范围为10—27种;6—12月的种类数波动较小,在11—17种之间变化(图1)。按照分类阶元划分,缘毛目纤毛虫所占比例最高,为35%;其次为侧口目纤毛虫,所占比例为12%,裸口目、管口目、毛口目和膜口目纤毛虫所占比例最低,均为2%(附表);在全年的各个月份,缘毛目纤毛虫的种类数所占比例均最高(图2)。
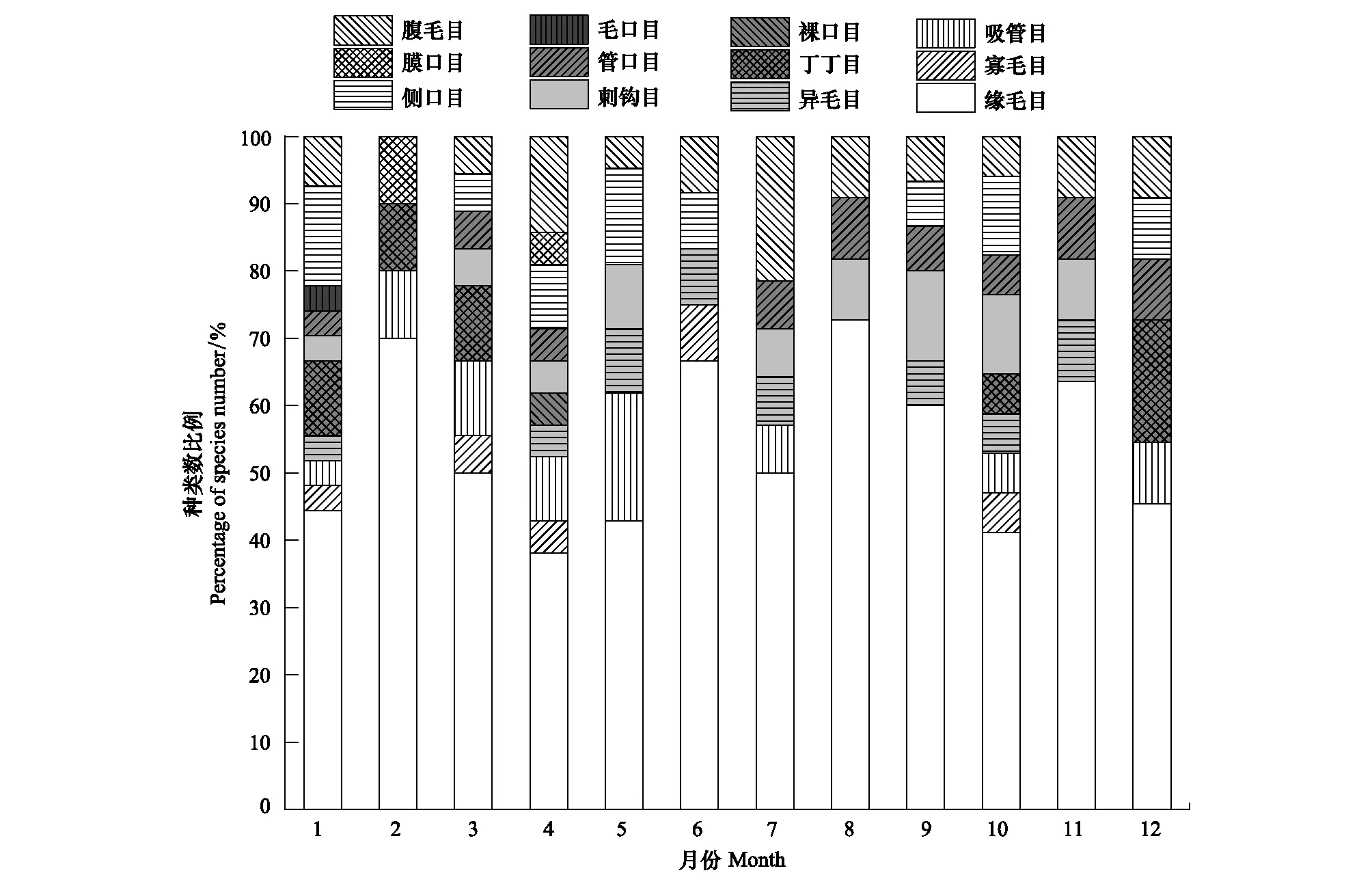
图2 周丛生纤毛虫各目的种类数所占比例的周年变化Fig.2 Annual variations of percentage of species number on different orders of periphytic ciliates
2.2.2 优势种密度的周年变化
沟钟虫(Vorticellaconvallaria),钟形钟虫(Vorticellacampanula),Vorticellainflusinum,钟虫sp. 2(Vorticellasp. 2),聚缩虫sp. 1(Zoothamniumsp. 1),螅状独缩虫(Carchesiumpolypinum),亨氏累枝虫(Epistylishentscheli)为优势种(表2)。沟钟虫密度在春季特别是5月最高,其次是在10—12月密度较高;钟形钟虫密度除在2月和6月以外的其他月份均较高;Vorticellainflusinum的密度在1月和3月出现两次峰值;钟虫sp. 2在6—8月密度较高,其他月份密度均较低;聚缩虫sp. 1在整个年度出现3次峰值,分别为3月,7月和10月,其他月份密度都较低;螅状独缩虫在12—4月的密度均较高,其他月份密度较低;亨氏累枝虫密度在整个年度出现两次峰值,分别为6月与9月,整体为5—9月密度最高,其他月份密度较低(图3)。

图3 周丛生纤毛虫优势种密度的周年变化Fig.3 Annual variations of abundance on dominant periphytic ciliates
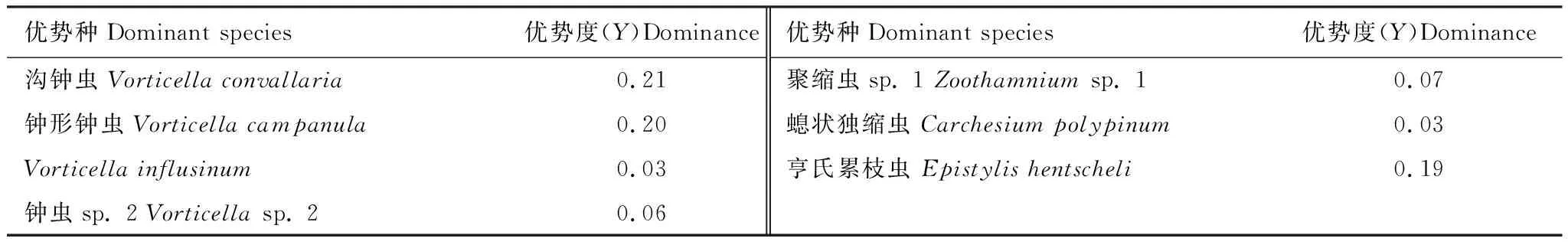
优势种Dominant species优势度(Y)Dominance优势种Dominant species优势度(Y)Dominance沟钟虫 Vorticella convallaria0.21聚缩虫sp. 1 Zoothamnium sp. 10.07钟形钟虫 Vorticella campanula0.20螅状独缩虫 Carchesium polypinum0.03Vorticella influsinum0.03亨氏累枝虫 Epistylis hentscheli0.19钟虫sp. 2 Vorticella sp. 20.06
2.3 周丛生纤毛虫多样性指数的周年变化
周丛生纤毛虫H′值从1—12月呈缓慢下降的趋势,变幅为1.05—2.10,年均值为1.53;D值变幅为1.91—6.33,年均值为3.16,变化情况与H′接近,且在8月最低,1月最高;J′值变幅为0.41—0.72,各月变化不大,年均值为0.56(图4)。
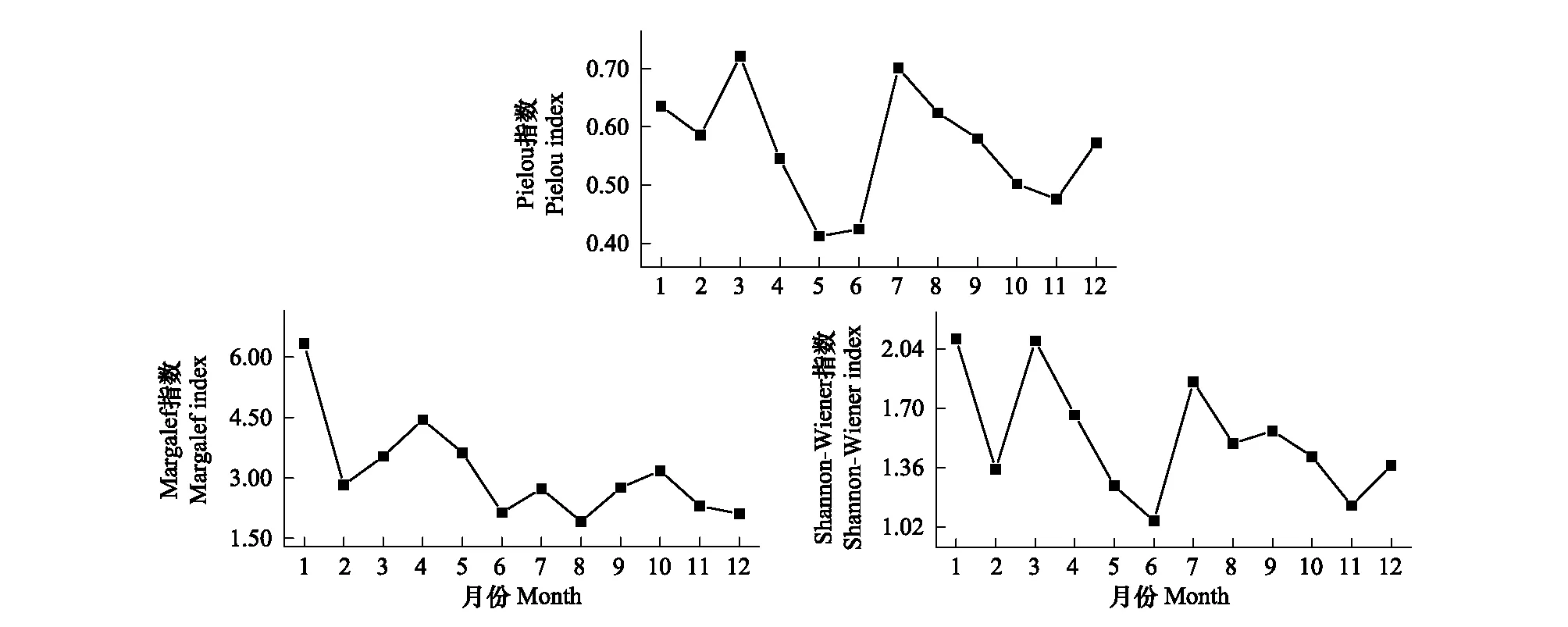
图4 周丛生纤毛虫多样性指数的周年变化Fig.4 Annual variations of diversity index of periphytic ciliates
2.4 周丛生纤毛虫群落结构的聚类分析
在相似系数为50%时,景观水体周丛生纤毛虫可分为4个类群(ANOSIM,P<0.001):第一类群(I)为在2月聚集的周丛生纤毛虫群落;第二类群(II)为在6月聚集的周丛生纤毛虫群落;第三类群(III)为在1月和3月聚集的周丛生纤毛虫群落;将其余月份聚集的周丛生纤毛虫群落归为第四类群(IV)。在相似系数为65%时,可将第四类群的周丛生纤毛虫分为四类:第一类(IVa)为在7,8和9月聚集的周丛生纤毛虫群落;第二类(IVb)为在5月聚集的周丛生纤毛虫群落;第三类(IVc)为在4月聚集的周丛生纤毛虫群落;第四类(IVd)为在10,11月和12月聚集的周丛生纤毛虫群落(图5)。
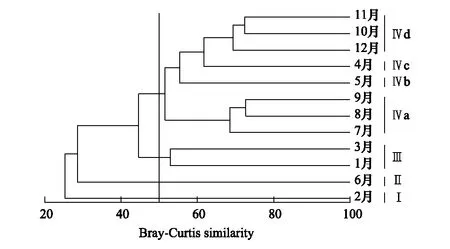
图5 周丛生纤毛虫群落的聚类分析树状图Fig.5 Clustering dendrogram of the periphytic ciliates communities
2.5 周丛生纤毛虫与环境因子的关系
2.5.1 周丛生纤毛虫群落结构参数与环境因子的相关性分析
对周丛生纤毛虫群落结构参数与水温(T)、溶解氧(DO)以及电导率(Spc)等11个水质参数进行Pearson相关性分析,周丛生纤毛虫的月平均密度与水体温度呈极显著正相关(P<0.01),与水体电导率呈显著正相关(P<0.05);丰富度指数与水体透明度呈极显著负相关(P<0.01)(表3)。对周丛生纤毛虫各个种的密度与环境因子进行BIOENV分析,在生物—环境相关性排序前10的环境变量组合中,总磷(TP)浓度和水温(T)是影响周丛生纤毛虫群落的主要环境因子,其中,周丛生纤毛虫的群落结构与TP、T、DO、pH(ρ值:0.625,P:0.001),TP、SD、T、DO、pH(ρ值:0.619,P:0.001)这两个环境因子组合的相关性最高(表4)。
表3周丛生纤毛虫的群落结构参数与环境因子的周年变动间Pearson相关性总结
Table3Pearsoncorrelationsbetweencommunitystructuralparametersofperiphyticciliatesandenvironmentalfactorswithannualvariations
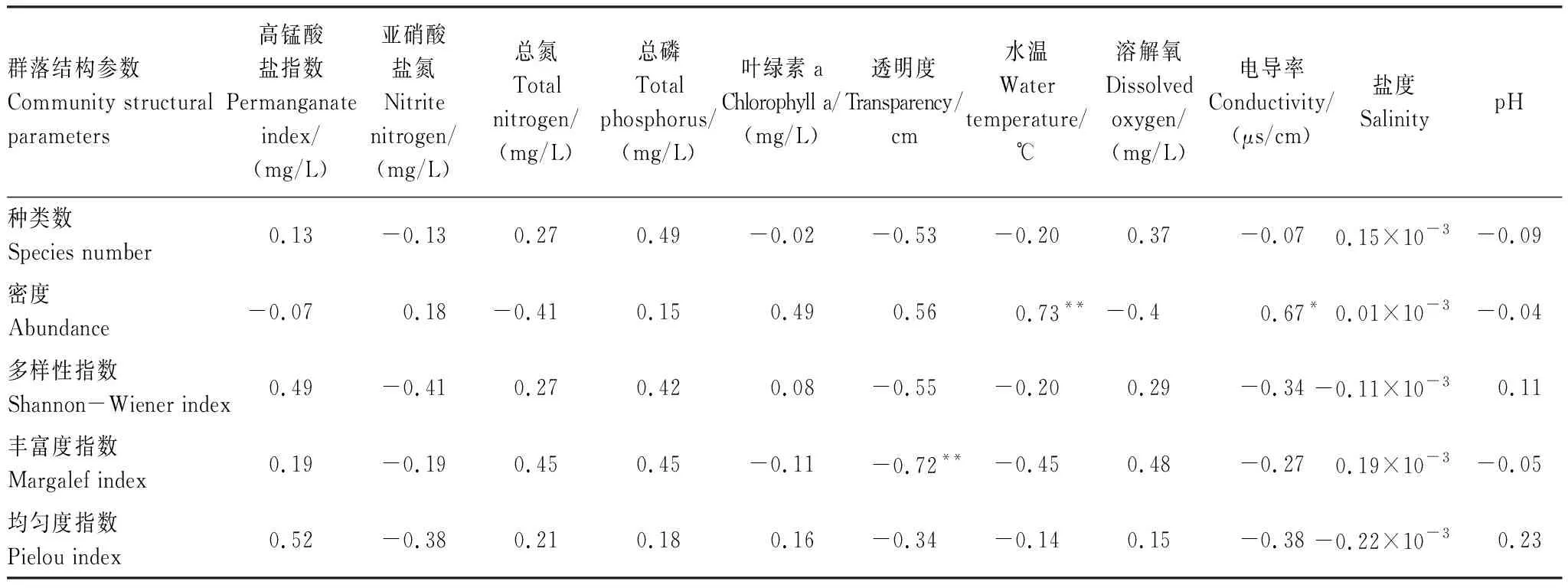
群落结构参数Community structural parameters高锰酸盐指数Permanganate index/(mg/L)亚硝酸盐氮Nitrite nitrogen/(mg/L)总氮Total nitrogen/(mg/L)总磷Total phosphorus/(mg/L)叶绿素aChlorophyll a/(mg/L)透明度Transparency/cm水温Water temperature/℃溶解氧Dissolved oxygen/(mg/L)电导率Conductivity/(μs/cm)盐度SalinitypH种类数Species number0.13-0.130.270.49-0.02-0.53-0.200.37-0.070.15×10-3-0.09密度Abundance-0.070.18-0.410.150.490.560.73**-0.40.67*0.01×10-3-0.04多样性指数Shannon-Wiener index0.49-0.410.270.420.08-0.55-0.200.29-0.34-0.11×10-30.11丰富度指数Margalef index0.19-0.190.450.45-0.11-0.72**-0.450.48-0.270.19×10-3-0.05均匀度指数Pielou index0.52-0.380.210.180.16-0.34-0.140.15-0.38-0.22×10-30.23
* 在0.05水平(双侧)上显著相关;** 在0.01水平(双侧)上显著相关
表4周丛生纤毛虫的群落与环境变量的生物-环境(BIOENV)分析总结
Table4Summaryofresultsfrombiota-environment(BIOENV)analysisshowingtheenvironmentalvariableswithtemporalvariationsintheperiphyticciliateabundances
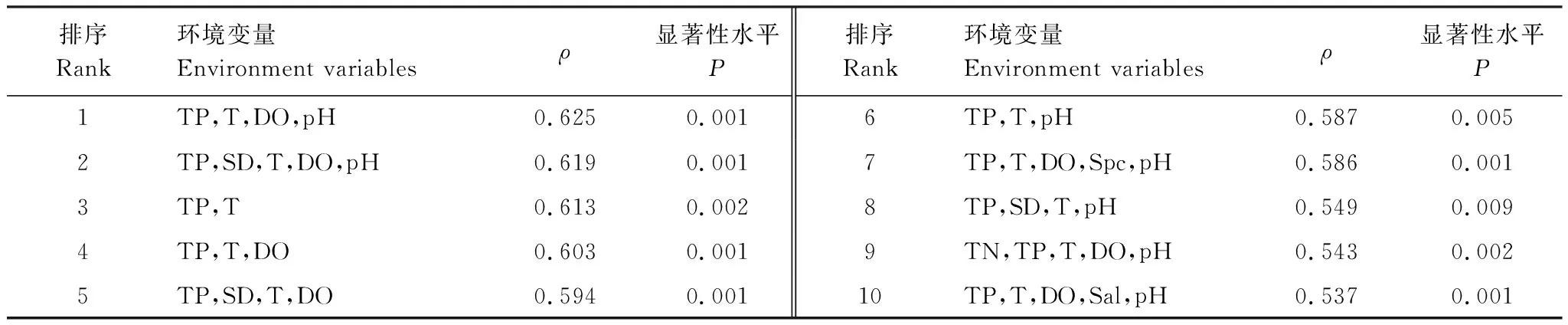
排序Rank环境变量Environment variablesρ显著性水平P排序Rank环境变量Environment variablesρ显著性水平P1TP,T,DO,pH0.6250.0016TP,T,pH0.5870.0052TP,SD,T,DO,pH0.6190.0017TP,T,DO,Spc,pH0.5860.0013TP,T0.6130.0028TP,SD,T,pH0.5490.0094TP,T,DO0.6030.0019TN,TP,T,DO,pH0.5430.0025TP,SD,T,DO0.5940.00110TP,T,DO,Sal,pH0.5370.001
TP:总磷,Total phosphorus;T:水温,Water temperature;DO:溶解氧,Dissolved oxygen;pH;SD:透明度,Transparency;Spc:电导率,Conductivity;TN:总氮,Total nitrogen;Sal:盐度,Salinity
2.5.2 周丛生纤毛虫各目的密度与水体环境因子之间的关系
Pearson相关性分析结果表明,缘毛目纤毛虫密度与透明度以及电导率呈显著正相关(P<0.05),与水温呈极显著正相关(P<0.01);丁丁目、毛口目和侧口目纤毛虫密度与透明度呈显著负相关(P<0.05),其中丁丁目纤毛虫密度与水温呈极显著负相关(P<0.01)、与总氮浓度呈极显著正相关(P<0.01),毛口目纤毛虫密度与总氮和总磷浓度均呈显著正相关(P<0.05)(表5)。
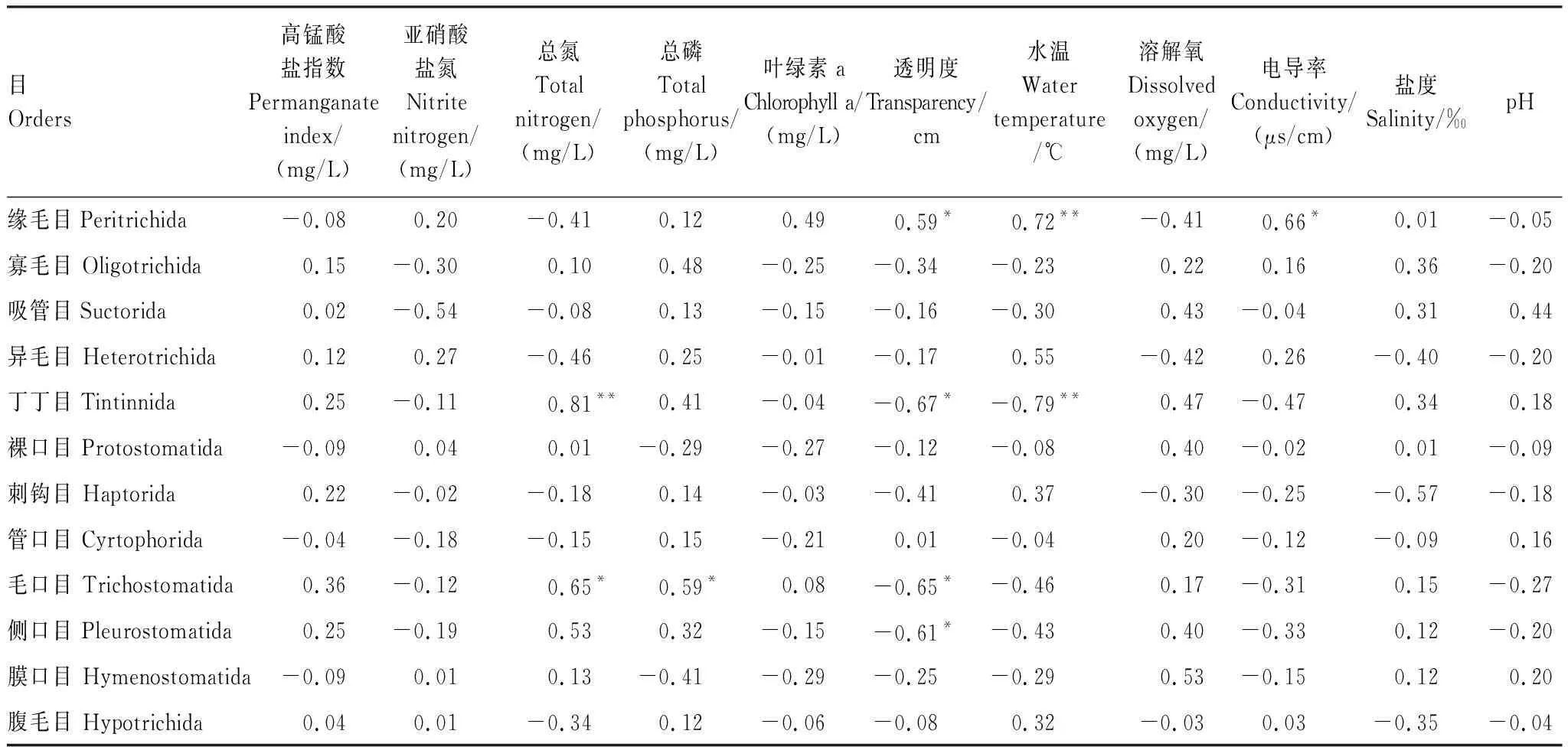
表5 周丛生纤毛虫各目的密度与环境因子的周年变动间Pearson相关性总结
* 在0.05水平(双侧)上显著相关;** 在0.01水平(双侧)上显著相关
2.5.3 周丛生纤毛虫优势种与水体环境因子之间的关系
对不同优势种的周从生纤毛虫密度与环境因子进行冗余分析(RDA)。前两个排序轴的特征值分别为0.42和0.29,第一排序轴解释了样本中42%的变异,第二排序轴解释了样本29%的变异,两者合并解释了样本71.10%的总变异(表6)。T、SD、DO、TN、TP、CODMn和Sal对周丛生纤毛虫各个优势种的影响较大(图6)。其中,钟形钟虫、钟虫sp. 2、聚缩虫sp. 1、亨氏累枝虫与SD和T呈正相关,与DO、TN和Sal呈显著负相关;沟钟虫与SD呈显著正相关,与TN、TP、CODMn和Sal呈显著负相关;螅状独缩虫与T、SD呈显著负相关,与DO、TN和Sal呈显著正相关。
表6周丛生纤毛虫优势种与环境因子的冗余分析统计特征
Table6Statisticalcharacteristicsofredundancyanalysisbasedonrelationshipsbetweendominantperiphyticciliatesandenvironmentalfactors
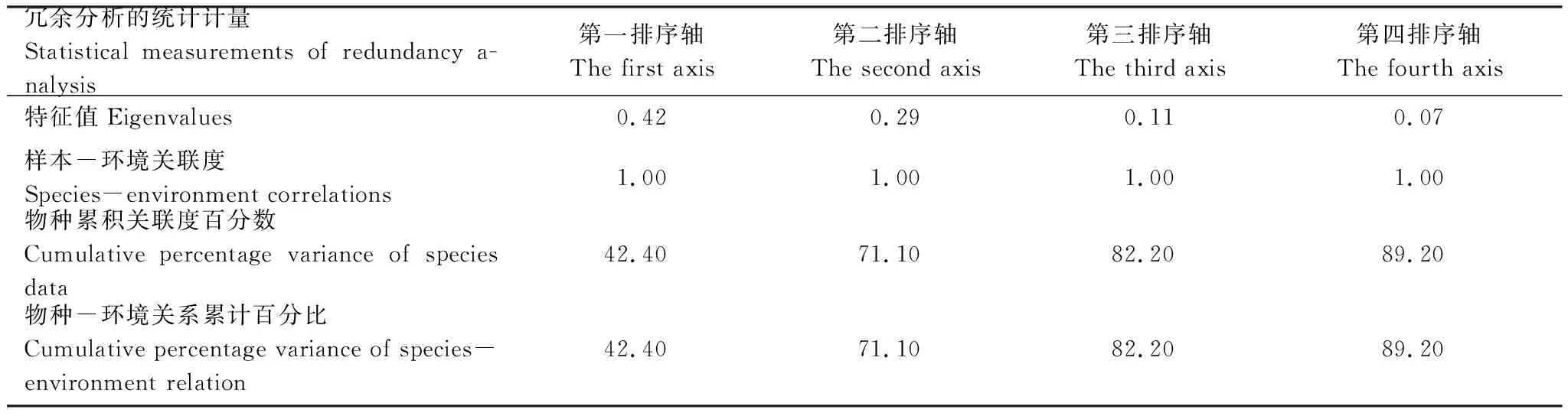
冗余分析的统计计量Statistical measurements of redundancy a-nalysis第一排序轴The first axis第二排序轴The second axis第三排序轴The third axis第四排序轴The fourth axis特征值Eigenvalues0.420.290.110.07样本-环境关联度Species-environment correlations1.001.001.001.00物种累积关联度百分数Cumulative percentage variance of species data42.4071.1082.2089.20物种-环境关系累计百分比Cumulative percentage variance of species-environment relation42.4071.1082.2089.20

图6 周丛生纤毛虫优势种与环境因子的冗余分析Fig.6 Redundancy analysis based on relationships between dominant periphytic ciliates and environmental factorsV. con:沟钟虫,Vorticella convallaria;V. cam:钟形钟虫,Vorticella campanula;V. inf:Vorticella influsinum;V. sp. 2:钟虫sp. 2,Vorticella sp. 2;Z. sp. 1:聚缩虫sp. 1,Zoothamnium sp. 1;C. pol:螅状独缩虫,Carchesium polypinum;E. hen:亨氏累枝虫,Epistylis hentscheli
3 讨论
3.1 周丛生纤毛虫的种类组成
周丛生纤毛虫的群落结构在沿海水域、海湾及湖泊水体中均具有季节性差异,其应对水体环境变化也呈现显著的季节性变动[11, 35]。Gong等[13]在胶州湾的周年研究中共检测出周丛生纤毛虫37种(30属)。Xu等[21]在韩国沿海水体中检测出周丛生纤毛虫29种。本研究周丛生纤毛虫(51种)的物种多样性显著高于上述近海岸的开放水体,这可能是与近海岸的开放水体相比,景观水体中的周丛生纤毛虫种类不易受潮汐扰动有关。而本研究所检测出的周丛生纤毛虫物种数显著低于Agamaliev[36]在里海水中检测出的周丛生纤毛虫物种数(130种),这可能与本研究的采样范围较小导致物种多样性较低有关。此外,本研究结果表明,该景观水体中的周丛生纤毛虫的群落结构存在明显的季节性差异,其中,冬季1月检测的周丛生纤毛虫种类最多,共27种;冬季2月种类数最少,共10种;周丛生纤毛虫以缘毛目纤毛虫为优势类群,其密度与种类数在全年的各个月份均占绝对优势,且全年的优势种均隶属于缘毛目,这可能与以载玻片作为人工基质、水体的水质状态以及食物较适合其生长有关。
3.2 周丛生纤毛虫群落结构与环境因子的关系
水体环境因子(如溶解氧、水温、硝酸盐氮、亚硝酸盐氮、可溶性磷酸盐、氨氮、盐度等)的变动是影响周丛生纤毛虫群落结构分布的重要因素。已有研究表明,海水养殖水体中周丛生纤毛虫的丰度和物种数量与可溶性磷酸盐具有极显著正相关关系[37],周丛生纤毛虫的密度与水体硝酸盐(NO3-N)和总磷(TP)的浓度呈正相关[38]。本研究的BIOENV分析也发现类似的结果,总磷(TP)浓度和水温(T)是影响周丛生纤毛虫密度的主要环境因子(表4)。
缘毛目纤毛虫的密度与温度和透明度呈极显著或显著正相关关系(表5)。可能因为采样水温为7.60—29.60℃,原生动物的种群增长率在该温度范围内随温度的升高而升高[39];同时它们为滤食性种类,主要以微微型浮游生物为食[40],通过其捕食能够提高水体透明度。本研究中,漫游类的侧口目纤毛虫密度与水体透明度呈显著负相关,这表明采集的侧口目纤毛虫更适宜于生存在透明度较低的水体中。虽然毛口目纤毛虫也与环境因子显著相关(与总氮浓度及总磷浓度呈显著正相关,与透明度呈显著负相关),但因仅发现一种Plagiopylasp.且只在1月检出,其相关性分析结果说服力较差。
不同种类最适温度的范围各异,如一些丁丁目的周丛生纤毛虫主要喜低温[41]。本研究中,丁丁目周丛生纤毛虫密度与水温呈极显著负相关(P<0.01),丁丁目的周丛生纤毛虫的主要种类如淡水筒壳虫(Tintinnidiumfluviatile)和恩茨筒壳虫(Tintinnidiumentzii)主要出现在温度较低的冬季,可能是由于这类周丛生纤毛虫喜低温,同时一些固着类的吸管目、缘毛目周丛生纤毛虫繁殖速度变缓,使得载玻片上的物种间的空间竞争速度减慢,丁丁目纤毛虫为代表的浮游类纤毛虫竞争压力变小,密度增大,从而也为该类物种在载玻片上提供了一个生存所需的空间。
3.3 优势种对水质的指示作用
在沿海水域、淡水及半咸水水体中,周丛生纤毛虫优势种多为缘毛目纤毛虫,其对不同类型的水体均具有一定的指示作用。如在青岛沿海的优势种聚缩虫未定种(Zoothamniumsp.)与pH、溶解氧以及营养盐显著相关[35];武汉东湖的优势种钟形钟虫与水温具有相关性;而青岛胶州湾的优势种中国伪钟虫(Pseudovorticellasinensis)与较低的溶解氧(3.80 mg/L)有关[13]。本研究的RDA分析表明,优势种的密度与水体的环境因子具有较大的相关性(图6)。其中,钟形钟虫、钟虫sp. 2、聚缩虫sp. 1、亨氏累枝虫、沟钟虫和螅状独缩虫这6个种的存在以及是否成为优势类群均可以考虑用作评定水体的质量状态,并且可以作为水体富营养化以及有机污染的潜在指示生物。但这一结论仍需进一步的实验进行验证。
综上所述,本研究发现载玻片法采集的周丛生纤毛虫原生动物同样也能反映景观水体水质。由于景观水体有面积小和水浅的特点,PFU上的小孔极易被搅起的底质或大颗粒悬浮物堵塞而造成群落结构失真无法正确监测水质,载玻片法可作为PFU法的一个良好补充或替代。
致谢:感谢上海海洋大学许朋和邢宏伟同学在样品采集和理化因子测定方面提供的帮助。
猜你喜欢
生态学报(2022年9期)2022-05-26 10:03:04
河南畜牧兽医(2021年1期)2021-01-07 11:38:30
河南畜牧兽医(2021年5期)2021-01-06 11:43:41
中国卫生标准管理(2020年18期)2020-10-16 09:32:36
江河文学(2020年6期)2020-01-04 07:06:37
渔业科学进展(2018年1期)2018-04-03 08:45:38
临床与实验病理学杂志(2017年12期)2017-03-20 03:48:46
浙江农业科学(2016年11期)2016-05-04 04:16:48
海洋与湖沼(2016年1期)2016-01-15 03:52:09
西藏科技(2015年6期)2015-09-26 12:12:08
