
Cr6+和Ni2+对拟康宁木霉的毒性及其抗性机制
2018-09-12 08:49林锦美段金明葛晶晶陈锦芳占易苍
集美大学学报(自然科学版) 2018年4期
林锦美,段金明,葛晶晶,陈锦芳,占易苍
(1.集美大学食品与生物工程学院,福建 厦门 361021;2.青岛市环境保护宣传教育中心,山东 青岛 266003;3.厦门三达膜环境股份有限公司,福建 厦门 361021)
0 引言
近年来,随着我国工业的快速发展,越来越多的重金属废水被排放出来[1]。重金属废水大部分具有毒性,且具有持久性、累积性,排放到环境水体中会导致生态系统的严重破坏,对环境和人类健康造成极大危害[2-4]。自然界中存在一些微生物,在高浓度金属条件下,仍能存活或生长,表现出对金属的抗性。有些菌还能通过生物转化作用或生理代谢活动使金属由高毒状态变为低毒状态,这为微生物法处理重金属废水提供了有利条件。相对于传统方法,微生物法具有环保、成本低、无二次污染等优势,在重金属废水治理中具有良好的发展前景[5-6]。
微生物对重金属的抗性机制主要包括胞外沉淀、细胞积累、解毒、生物转化等[7-10]。由于重金属是氧供体的一种强竞争离子,具有强氧化特性,在很多生物中都能诱发产生过氧化氢(H2O2)、超氧阴离子(O2-·)、氢氧自由基(·OH)等活性氧自由基(reactive oxygen species,ROS),对细胞内的膜质、蛋白质、DNA 等有很高的反应活性和损伤作用。微生物对重金属有抗性是因为体内存在抗氧化系统。生物系统的抗氧化系统包括抗氧化酶和抗氧化剂。抗氧化酶主要有超氧化物岐化酶(superoxide dismutase,SOD)和过氧化氢酶(catalase,CAT);抗氧化剂主要有胞内的还原型谷胱甘肽(glutathione,GSH)。因此,SOD和GSH可以作为微生物抗重金属强弱的标准。目前,木霉属处理含有重金属的废水研究已有很多报道[11-12],主要包括曲霉属(Aspergillus)、根霉属(Rhizopus)、青霉菌(Penicillium)等,且这些报道主要集中在对重金属的吸附率、影响吸附率的因素等方面,而对于木霉属在处理含重金属废水,尤其对重金属抗性性能的研究鲜有报道[11-12]。
本文运用自行筛选、培养、驯化及初步鉴定的拟康宁木霉(Trichodermakoningiopsis),对Cr6+和Ni2+的抗性机制进行初步探索,研究不同重金属离子对木霉的抗氧化系统及菌量的影响,同时就不同浓度外源NO和SO2对重金属作用下菌体的抗氧化系统的影响进行研究,为阐明NO/SO2影响重金属胁迫下菌体生长的生理生化机理提供理论依据。
1 材料与方法
1.1 实验材料
1.1.1 菌种和培养基 拟康宁木霉由集美大学环境工程实验室分离鉴定并保存。培养基的制备:实验采用配好的土豆培养基(m(土豆)∶m(水)=1∶5,葡萄糖25.4 g/L,KH2PO42.1 g/L,MgSO42.1 g/L)灭菌后接种,30 ℃培养3 d。
1.1.2 模拟废水 Ni2+标准储备液:准确称取49.32 g Ni(NO3)·6H2O,用蒸馏水溶解,稀释定容至1 000 mL,配制质量浓度为10 g/L含Ni2+的标准贮备液,于棕色试剂瓶内避光保存。Cr6+标准储备液:准确称取28.28 g K2Cr2O7,用蒸馏水溶解,稀释定容至1 000 mL,配制质量浓度为10 g/L含Cr6+的标准贮备液,于棕色试剂瓶内避光保存。本试验所用铜溶液均由贮备液稀释得到。
1.2 仪器和试剂
紫外可见分光光度计(UV751GD,上海精密仪器仪表有限公司),恒温振荡器(SHA-C,江苏省金坛市鸿科仪器厂),智能生化培养箱(PHX,宁波莱福科技有限公司),台式高速离心机(H1650,湖南湘仪实验室仪器开发有限公司)。
重铬酸钾(优级纯),硝酸镍(优级纯),硝酸(分析纯),氢氧化钠(分析纯)。低粘度树脂包埋试剂盒Spurr Embedding Kit(北京海德公司)。
1.3 实验方法
1.3.1 毒性及抗性实验 将具有抗Cr6+和Ni2+菌株活化24 h后,分别在不同Cr6+、Ni2+浓度的条件下,用接种环将驯化后的木霉菌种接到液体培养基中,于30 ℃、转速为120 r/min培养箱里恒温振荡培养3 d,然后将培养后的菌液置于抽滤机内以0.05 MPa压力抽滤,并用去离子水反复清洗,最后将得到的菌种置于65 ℃烘箱内24 h烘干,用电子天平称其干重以表示细菌的生长量,并测定获得的菌粉中GSH和SOD含量。
1.3.2 GSH提取及其测定 参照文献[13]采用热水抽提法。取1 g烘干的菌粉,与4 mL蒸馏水充分混合后倒入5 mL沸腾的水中,充分混合后放入95~100 ℃水浴锅中水浴10 min后,立即取出放于冰水中速冷,然后在转速为5 000 r/min下离心10 min,取出上清液,用碘量法测定GSH的含量。
1.3.3 SOD提取及测定 每毫克组织蛋白质在1 mL反应液中SOD抑制率达 50%时所对应的SOD量作为一个SOD活力单位(U)。培养后的菌液经离心沉淀后用考马斯亮兰法(蛋白质试剂盒,南京建成生物工程研究所)测定出组织中蛋白质含量。每个体系取3个试样作为平行实验,并计算酶液的可溶性蛋白质含量。
1.4 数据处理
每组实验均重复3次,取平均值,并运用Origin软件进行数据处理与图表绘制。采用Excel进行方差分析,显著性水平为0.05。
2 结果
2.1 Cr6+和Ni2+对拟康宁木霉的毒性
在含有不同重金属溶液中接种拟康宁木霉进行毒性分析试验,结果如图1所示。由图1可见,在重金属Cr6+和Ni2+的胁迫下,菌体的生长均受到严重影响,2种重金属对拟康宁木霉的生长始终表现出抑制作用。说明重金属对菌体有一定毒性作用,且Cr6+对菌体的毒性强于Ni2+。当Cr6+质量浓度从0升高到150 mg/L时,拟康宁木霉的生长量显著下降,同时,菌体内GSH的含量呈现先增加后降低的趋势,并且在Cr6+的质量浓度为100 mg/L时达到最高。Ni2+对拟康宁木霉的影响跟Cr6+相似,当Ni2+质量浓度从0升高到300 mg/L时,菌体的生长量迅速降低,而菌体内GSH的含量先增加后减少,且在Ni2+质量浓度为100 mg/L时,GSH的含量达到最高。由此可知,重金属的存在能够诱导拟康宁木霉体内产生GSH来减小对其的毒性作用。

2.2 外源NO对Cr6+和Ni2+胁迫下拟康宁木霉抗性
2.2.1 外源NO对Cr6+和Ni2+胁迫下拟康宁木霉生长量的影响
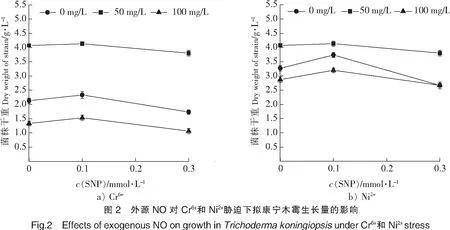
本实验通过测定NO的供体硝普钠(sodium nitroprusside,SNP)[14]的浓度来反映NO对Cr6+和Ni2+胁迫下拟康宁木霉生长的影响,结果见图2。由图2可知,在没有Cr6+和Ni2+时,SNP浓度由0增加到0.1 mmol/L时,拟康宁木霉的生长量没有变化;SNP浓度由0.1 mmol/L增加到0.3 mmol/L时,拟康宁木霉的生长有所抑制。当Cr6+和Ni2+质量浓度增加到50 mg/L时,Cr6+和Ni2+对拟康宁木霉的生长均呈现一定的抑制作用,但是加入0.1 mmol/L的SNP,菌体的生长量反而增加,SNP增加到0.3 mmol/L时,菌体的生长量有所降低。当Cr6+和Ni2+质量浓度为150 mg/L时,结果类似。说明一定浓度的NO能够降低Cr6+和Ni2+对菌体的毒性作用,促进菌体的生长,但NO浓度过高,却不利于菌体的生长。
2.2.2 外源NO对Cr6+和Ni2+胁迫下拟康宁木霉GSH含量的影响
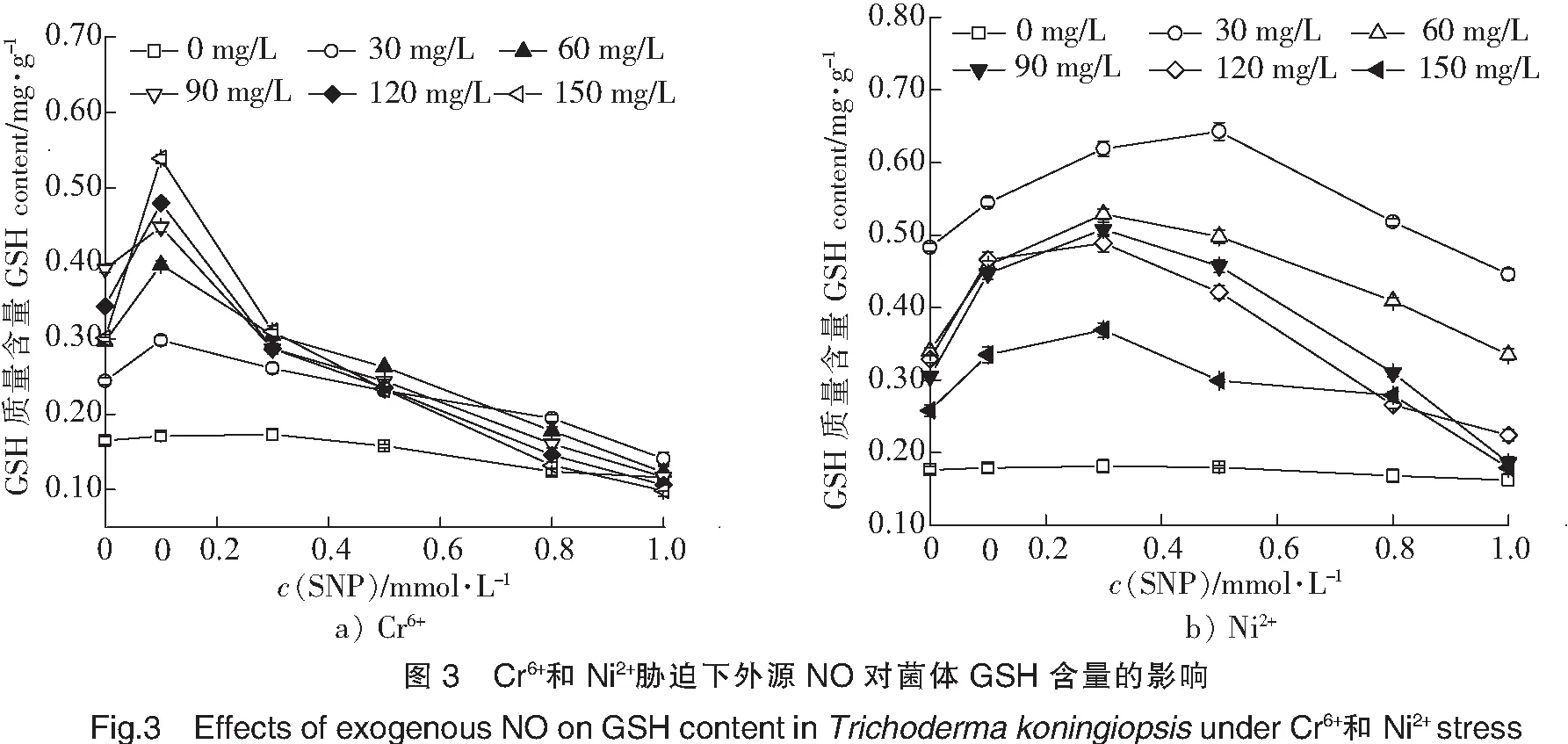
本实验研究了不同外源NO浓度对拟康宁木霉中GSH含量的影响,结果如图3所示。由图3可以看出,在含有Cr6+的条件下,当SNP浓度由0升高到0.2 mmol/L时,菌体内GSH含量也随之升高。继续升高SNP浓度,菌体内GSH的含量迅速降低。由此表明,一定浓度的外源NO能够促进菌体内GSH含量的增加,从而抵抗菌体对Cr6+和Ni2+的毒性。在Ni2+的条件下,结果和Cr6+的情况类似,当SNP浓度由0升高到0.3 mmol/L时,菌体内GSH含量也随之升高,继续升高SNP浓度,菌体内GSH的含量降低。这说明,一定浓度的外源NO能够促进菌体内GSH含量的增加,从而抵抗菌体对Cr6+和Ni2+的毒性,但是NO浓度太高时,反而会起抑制作用。
2.2.3 外源NO对Cr6+和Ni2+胁迫下拟康宁木霉SOD活性的影响
本实验研究了不同浓度的外源NO对拟康宁木霉SOD活性的影响,结果如图4所示。从图4中可以看出,在不存在重金属Cr6+和Ni2+的条件下,菌体中SOD的活性很低。这是因为没有重金属的诱导,不会激发菌体中SOD的活性来抵抗重金属的毒性。在没有外源NO的加入时,随着Cr6+和Ni2+浓度的增加,菌体内SOD的活性呈现先增加后降低的趋势,且重金属Cr6+和Ni2+质量浓度分别为120,150 mg/L时,菌体内SOD活性最高。这说明一定浓度重金属,可以激发菌体中SOD的活性来抵抗重金属的毒性,但是重金属浓度太高,对菌体的毒害作用太大,所以菌体内SOD的活性反而会降低。当加入外源NO时,菌体中SOD的活性随着外源NO浓度的升高呈现先增强后降低的趋势,且在NO浓度为0.5 mmol/L时,SOD活性最高。这表明,低浓度外源NO可以促进菌体内SOD活性增强,而高浓度的外源NO对菌体内SOD活性却存在抑制作用。

2.3 外源SO2对Cr6+和Ni2+胁迫下拟康宁木霉抗性
本实验分别研究了在Cr6+和Ni2+初始质量浓度为0,50,100 mg/L时不同浓度外源SO2对菌体内GSH含量及SOD活性的影响,结果见图5。由图5可以看出,在Cr6+胁迫的菌体中,当外源SO2浓度由0增加到0.5 mmol/L时,GSH的含量及SOD的活性迅速降低,而随着外源SO2浓度由0.5 mmol/L升高到1.0 mmol/L时,菌体内GSH的含量下降速度有所减缓,说明SO2对菌体中GSH合成及SOD活性产生抑制作用。可能原因是:外源SO2在菌体上发生了氧化反应,此过程产生了大量的活性氧,加速了菌体的氧化损伤,抑制了菌体内GSH的合成及SOD的活性。

2.4 Cr6+和Ni2+对拟康宁木霉的毒性和抗性相关性分析
通过对不同浓度Cr6+和Ni2+对拟康宁木霉的毒性和抗性相关性分析表明,不同浓度重金属胁迫下, Cr6+毒性和抗性相关系数为-0.758 8(P<0.05),表现出显著负相关;而Ni2+毒性与抗性相关系数为-0.298 7,其负相关性不明显。外源NO条件下Cr6+和Ni2+对拟康宁木霉抗性相关性分析结果见表1。由表1可知,在不同Cr6+和Ni2+浓度胁迫下,低浓度外源NO中的GSH与SOD表现出较显著的正相关性(P<0.05),高浓度外源NO中的GSH与SOD相关性不显著。

表1 外源NO对不同浓度Cr6+和Ni2+胁迫下拟康宁木霉抗性GSH/SOD相关性
3 讨论
关于木霉处理废水的研究已有一些报道,如李会东等用木霉处理含铬废水,在28 ℃、pH值为1的环境条件下,其对Cr6+的吸附率高达99%[15]。但这些研究大多集中在生物吸附方面,而对于其抗重金属机制的研究并没有深入进行。本研究发现,拟康宁木霉在处理重金属废水时体内GSH的含量及SOD的活性发生显著的变化,在Cr6+和Ni2+胁迫下,GSH的含量及SOD的活性明显高于无Cr6+和Ni2+胁迫下的。当存在Cr6+和Ni2+时,Cr6+和Ni2+诱导菌体产生GSH和增强SOD的活性,从而抵抗重金属的毒性,这跟陈叶福等[16]的研究结果类似。然而,在重金属诱导下,拟康宁木霉是通过什么途径合成GSH并提高SOD的活性的,本文并没有深入研究。从分子和代谢水平阐明GSH的合成机制和途径,可能成为未来的研究重点。另外,高浓度Cr6+和Ni2+对拟康宁木霉有明显的抑制作用,而实际中有一些工业废水如电镀废水、冶金废水等,其重金属离子浓度极高,这对微生物处理提出新的挑战。因此,如何提高微生物对重金属离子的抗性,就显得尤为迫切。也许通过基因工程构建工程菌,为我们提供了一条可行的途径。
NO是一种广泛分布于生物体内的气体活性分子。有研究表明[17],外源NO能够提高盐胁迫下种子的萌发,延缓果实等器官的成熟和衰老,以及参与植物抗病反应等,在植物体内对各种生物和非生物胁迫反应的信息传递发挥着重要的作用。但是外源NO浓度过高对生物体也具有毒害作用,NO对生物体的代谢具有双重性[17-19]。而NO对微生物抗重金属胁迫的研究,目前鲜有报道。本文研究了不同外源NO浓度对拟康宁木霉抗重金属的影响,结果显示,低浓度NO有利于菌体内GSH的合成及SOD活性的提高,从而抵抗重金属的毒性,而高浓度外源NO反而有抑制作用。这说明,NO不仅能够调节植物对重金属胁迫下的抗性强弱,对微生物也有相似的作用。但是关于NO如何调节微生物的抗性,目前仍不清楚。本实验从NO调节菌体中GSH的合成及SOD的活性来初步阐述了NO对微生物抗重金属的机制,但是具体的分子机制还有待进一步研究。
4 结论
1)在Cr6+和Ni2+的作用下,随着重金属浓度增加,拟康宁木霉菌生长量逐渐减少,GSH含量先增加后减少。说明重金属诱导菌体生成GSH,缓解重金属对细胞的氧化损伤作用,而Cr6+胁迫下,菌量与GSH呈现显著的负相关性,Ni2+胁迫下,菌量与GSH的负相关性不显著。重金属Cr6+对木霉的毒性作用强于Ni2+。
2)低浓度外源NO能够增加菌体内GSH含量及SOD的活性,从而缓解重金属胁迫对菌体的氧化损伤作用,且二者呈现正相关性(P<0.05);高浓度外源NO则加重重金属对菌体的损伤作用,GSH/SOD相关性不显著。
3)在不同浓度外源SO2作用下,GSH的含量和SOD的活性均随外源SO2浓度的增加而降低,说明外源SO2加剧重金属对菌体的损伤作用。
猜你喜欢
当代水产(2022年1期)2022-04-26
内燃机工程(2021年5期)2021-10-19
干旱地区农业研究(2021年4期)2021-08-11
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
食品与发酵工业(2021年7期)2021-04-27
小学生作文辅导(2021年9期)2021-04-14
食品研究与开发(2021年1期)2021-01-21
江苏教育·书法教育(2021年3期)2021-01-17
中国调味品(2017年2期)2017-03-20
创新科技(2014年12期)2014-07-27
