
Enriched environment elevates expression of growth associated protein-43 in the substantia nigra of SAMP8 mice
2018-09-11 07:26:58ZhenYunYuanJieYangXiaoWeiMaYanYongWangMingWeiWang
中国神经再生研究(英文版) 2018年11期
Zhen-Yun Yuan , Jie Yang, Xiao-Wei Ma, Yan-Yong Wang, Ming-Wei Wang
The First Hospital of Hebei Medical University; Brain Aging and Cognitive Neuroscience Laboratory of Hebei Province, Shijiazhuang, Hebei Province,China
Abstract An enriched environment protects dopaminergic neurons from 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-induced neuronal injury, but the underlying mechanism for this is not clear. Growth associated protein-43 (GAP-43) is closely associated with neurite outgrowth and axon regeneration during neural development. We speculate that an enriched environment can reduce damage to dopaminergic neurons by affecting the expression of GAP-43. This study is designed to test this hypothesis. Three-month-old female senescence-accelerated mouse prone 8 (SAMP8) mice were housed for 3 months in an enriched environment or a standard environment. These mice were then subcutaneously injected in the abdomen with 14 mg/kg MPTP four times at 2-hour intervals. Morris water maze testing demonstrated that learning and memory abilities were better in the enriched environment group than in the standard environment group.Reverse-transcription polymerase chain reaction, immunohistochemistry and western blot assays showed that mRNA and protein levels of GAP-43 in the substantia nigra were higher after MPTP application in the enriched environment group compared with the standard environment group. These findings indicate that an enriched environment can increase GAP-43 expression in SAMP8 mice. The upregulation of GAP-43 may be a mechanism by which an enriched environment protects against MPTP-induced neuronal damage.
Key Words: nerve regeneration; Parkinson’s disease; neural plasticity; senescence-accelerated mouse prone 8; growth associated protein-43;substantia nigra; learning and memory; neural regeneration
Introduction
Parkinson’s disease (PD) is a common neurodegenerative disease in the elderly. Its main feature is gradual loss of dopaminergic neurons in the substantia nigra, accompanied by motor symptoms and cognitive impairments and sometimes dementia (Wirdefeldt et al., 2011). PD affects 1–2% of the population over the age of 60 years (Ohashi et al., 2006; Wood et al., 2010; Pringsheim et al., 2014). Motor impairments and cognitive dysfunction are usual symptoms of PD (Wood et al., 2010) that result from extended death of dopaminergic neurons in the substantia nigra pars compacta and a 70–80%reduction in dopamine levels in the striatum (Bergman and Deuschl, 2002; Galvan and Wichmann, 2008). PD is considered a multi-factor disorder that results from the combined effects of multiple factors, including genetic and environmental factors and aging (Elbaz et al., 2016), with aging being the most prominent risk factor (Collier et al., 2007).
The senescence-accelerated mouse prone 8 (SAMP8) inbred mouse line has early onset senility and a short life span(Flood and Morley, 1998) and is characterized by learning and memory impairments, and affective disturbance during the aging process (Kawamata et al., 1997; Takeda et al., 1997). In our previous studies, we have used a 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-SAMP8 model of PD (Liu et al., 2008; Yuan et al., 2008), which has non-motor symptoms (Liu et al., 2010).
With an aging population, the number of people with PD is increasing (LaHue et al., 2016). Although several studies have suggested that combinations of pharmacological and non-pharmacological therapies can alleviate PD symptoms(Mo et al., 2015; Chang et al., 2016), all therapeutic options to date have failed to halt the progression of degeneration(Kakkar and Dahiya, 2015). Some experimental data indicate that the environment can change behavior and molecular indicators in some animal models (Laviola et al., 2008).These changes can induce plastic brain changes and promote beneficial effects on the progression of neuronal impairment related to PD (Campêlo et al., 2017). Physical activity and experience of novelty have been shown to improve both motor and cognitive functions in PD patients (Benka Wallén et al., 2015; Hindle et al., 2016).
A previous study has shown that enriched environments(EEs) can protect dopaminergic neurons from MPTP-induced neuronal injury (Yuan et al., 2009a). What are the underlying mechanisms for this effect? In the past few years,many studies have focused on neural plasticity. Growth associated protein-43 (GAP-43) is strongly associated with neurite growth and axon regeneration during neural development (Aigner and Caroni, 1993). GAP-43 is also regarded as a molecular marker of neural plasticity (Gorup et al.,2015). To investigate the possible protective effects of EEs,this study examined the cognitive ability of MPTP-treated SAMP8 mice in the Morris water maze (MWM), and determined mRNA and protein levels of GAP-43 in the substantia nigra pars compacta.
Materials and Methods
Animals
Ninety-six 3-month-old female SAMP8 mice weighing 20–23 g were obtained from the Animal Center of Hebei Medical University of China (No. SCXK (Ji) 2008-1-003).All mice were randomly and equally assigned to EE and standard environment (SE) groups. The EE cages (52 cm ×37 cm × 22 cm) consisted of a platform, several tunnels, two running wheels, a variety of toys and nesting material. There were 10–12 mice per cage. The SE cages (32 cm × 20 cm × 15 cm) only contained nesting material and 5–6 mice per cage.Mice were housed in a room at 22–25°C with 55 ± 5% humidity and a 12-hour light/dark cycle. Mice had free access to water and food and these conditions were maintained for 3 months. All experimental procedures were performed according to the Guidance Suggestions for the Care and Use of Laboratory Animals, issued by the Ministry of Science and Technology of China. This study was approved by the Animal Ethics Committee of the First Hospital of Hebei Medical University of China (approval No. 20131001).
MPTP injection
Mice were equally and randomly divided into an MPTP group and a 0.9% normal saline solution (NS) group. The MPTP mice were subcutaneously injected in the abdomen with MPTP (14 mg/kg) four times from 8 am at 2-hour intervals. The mice in the NS group received an equal volume(14 mL/kg) of 0.9% saline solution at the same time point.MWM experiments were conducted for 5 days. The following day, mice were euthanized for immunohistochemistry,and reverse-transcription polymerase chain reaction (RTPCR) and western blot assays.
MWM test
The day after MPTP and NS injection, 96 mice were tested in the MWM for 5 consecutive days using apparatus from Anhui Huaibei Zhenghua Biological Equipment (Anhui,China). The swimming pool, 50 cm in height and 120 cm in diameter, had four quadrants. A hidden platform, 20 cm in height and 14 cm in diameter, was fixed 1.5 cm under the surface of the water and was defined as the target quadrant. The water temperature was maintained at 22 ± 2°C and was made opaque with black non-toxic dye to prevent the mice from seeing the platform. A place navigation test was performed for four days. Mice were put into the pool facing the wall of the pool and allowed to swim randomly until the platform was found or for up to 120 seconds. If a mouse could not find the platform, they were guided to the platform. All mice were allowed to stay on the platform for 10 seconds. Escape latency(the time taken to find the platform) was recorded. This was repeated three more times with the mice being placed in a different quadrant each time. On day 5, the probe test was performed; the hidden platform was removed and each mouse was allowed to swim for 120 seconds. The swim paths were automatically recorded with a video tracking system (Anhui Huaibei Zhenghua Biological Equipment) and the time spent in each quadrant was recorded.
RT-PCR
The day after the MWM test, eight mice from each group were euthanized. The substantia nigra was dissected on ice from each mouse. Total RNA was purified from fresh tissue,and cDNA was prepared for RT-PCR. β-Actin served as the internal reference. Primers were synthesized by Shanghai Bioasia Daily Chemical Co., Ltd. (Shanghai, China). The primers used are listed in Table 1.
For amplification of β-actin and GAP-43, the number of cycles was 27 and 30, respectively. Amplification conditions were as follows: predenaturation at 95°C for 5 minutes, 30 cycles of 95°C for 30 seconds, 54°C for 45 seconds and 72°C for 45 seconds, followed by a final extension at 72°C for 10 minutes. After electrophoresis, gel images were scanned anddata were analyzed with a JEDA 801E Gel image analyzer(Nanjing University Company). PCR products were quantitatively analyzed. Results are expressed as the ratio of the absorbance value of GAP-43 to that of β-actin.

Table 1 Primers for reverse-transcription polymerase chain reaction
Immunohistochemistry staining
The day after the MWM test, eight mice from each group were anesthetized with 10% chloral hydrate, perfused through the left ventricle with saline, and the brains were embedded in paraffin. According to The Mouse Brain Atlas in Stereotaxic Coordinates, the substantia nigra was sectioned in to consecutive 5-μm sections. Every fifth section was selected for immunostaining. Sections were incubated in 10%normal goat serum for 1 hour at 37°C and then incubated overnight at 4°C with rabbit anti-GAP-43 antibody (1:100;Chemicon Biotechnology, Temecula, CA, USA). Sections were then washed in 0.1 M PBS and incubated sequentially with biotinylated goat anti-rabbit IgG (1:100; Zhongshan Golden Bridge Biotechnology, Beijing, China) for 30 minutes at 37°C, horseradish peroxidase-conjugated streptavidin(1:100; Zhongshan Golden Bridge Biotechnology) for 30 minutes at 37°C, and then brie fly in diaminobenzidine. The cytoplasm of cells located in the substantia nigra pars compacta was positive for GAP-43, visible as a brownish yellow stain. Pictures were taken at 200× magnification using a BX53 microscope (Olympus, Tokyo, Japan) and analyzed using the Motic Med 6.0 digital medical image analysis system. Three sections were selected from each mouse, and a visual field on one side of the substantia nigra compacta was randomly selected from each section. The corrected optical density value of GAP43-ir was determined and the mean value obtained.
Western blotting
The day after the MWM test, eight mice of each group were euthanized. Protein from fresh substantia nigra tissue was extracted. For each sample, 60 µg of protein were subjected to polyacrylamide gel electrophoresis, and then transferred to a polyvinylidene fluoride membrane. Membranes were incubated with rabbit anti-GAP-43 polyclonal antibody(1:500; Chemicon Biotechnology, Temecula, CA, USA), or rabbit anti-β-actin polyclonal antibody (1:200; Zhongshan Golden Bridge Biotechnology) at room temperature for 1 hour, followed by horseradish peroxidase-conjugated goat anti-rabbit secondary antibody (1:1000; Santa Cruz Biotechnology, Santa Cruz, CA, USA) for 2 hours at room temperature. The products were developed with Super ECL Plus luminescence fluid (Applygen Technologies Inc., Beijing,China). Following film scanning, visualized bands were analyzed using Quantity One-4.6.2 software (Bio-rad, Hercules,CA, USA). β-Actin was used as the internal reference to confirm equal loading of the samples.
Statistical analysis
Data, presented as the mean ± SD, were analyzed with SPSS 11.5 software (SPSS, Chicago, IL, USA). The escape latency in the place navigation test of the MWM task was analyzed by two-way repeated measures analysis of variance followed by the least significant difference post hoc test. All other data analyses were performed by one-way analysis of variance followed by the least significance difference test. P values <0.05 were considered statistically significant.
Results
EE improved the learning and memory abilities of MPTP-treated SAMP8 mice Learning acquisition
Compared with the SE + NS group, escape latencies were significantly shorter in the EE + NS group (P < 0.01), but significantly longer in the SE + MPTP group (P < 0.01).Compared with the EE + NS group, escape latencies were longer in the EE + MPTP group (P < 0.01). Compared with the SE + MPTP group, escape latencies were shorter in the EE + MPTP group over four consecutive days (P < 0.01).The escape latency of each group shortened significantly with increasing number of trials, illustrating learning acquisition (Figure 1).
Memory retention
The percentage of time spent in the target quadrant was significantly longer (P < 0.001) and the percentage of time spent in the opposite quadrant was significantly shorter (P < 0.01)in the EE + NS group compared with the SE + NS group.After MPTP injection, the percentage of time spent in the target quadrant was significantly shorter (P < 0.001) and the percentage of time spent in the opposite quadrant was signi ficantly longer (P < 0.05) in the EE + MPTP group compared with the EE + NS group. Moreover, the percentage of time spent in the target quadrant was significantly shorter (P < 0.05)and the percentage of time spent in the opposite quadrant was significantly longer (P < 0.05) in the SE + MPTP group compared with the SE + NS group. However, compared with the SE + MPTP group, the percentage of time spent in the target quadrant was significantly longer (P < 0.05) and the percentage of time spent in the opposite quadrant was signi ficantly shorter (P < 0.05) in the EE + MPTP group (Figure 2).
Effect of EE on the levels of GAP-43 mRNA in the substantia nigra of MPTP-treated mice
Compared with the SE + NS group, GAP-43 mRNA levels were significantly increased in the EE + NS group (P < 0.05).MPTP resulted in a significant increase in GAP-43 mRNA levels both in the EE + MPTP group (P < 0.01) and the SE+ MPTP group (P < 0.01). Compared with the SE + MPTP group, mRNA levels of GAP-43 were higher in the EE +MPTP group (P < 0.01; Figure 3).
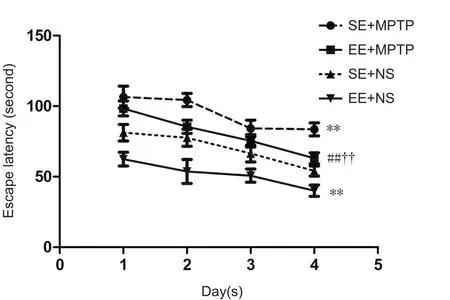
Figure 1 Effects of EE and MPTP on the average escape latency of SAMP8 mice in the place navigation test of the Morris water maze task.
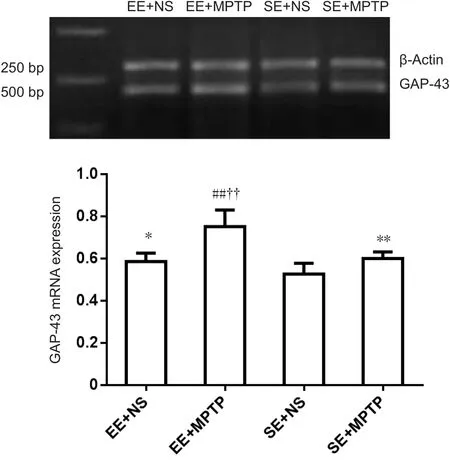
Figure 3 Effects of EE and MPTP on the relative level of GAP-43 mRNA in the substantia nigra of SAMP8 mice.
Effect of EE on GAP-43 immunoreactivity (GAP-43-ir) in the substantia nigra of MPTP-treated mice
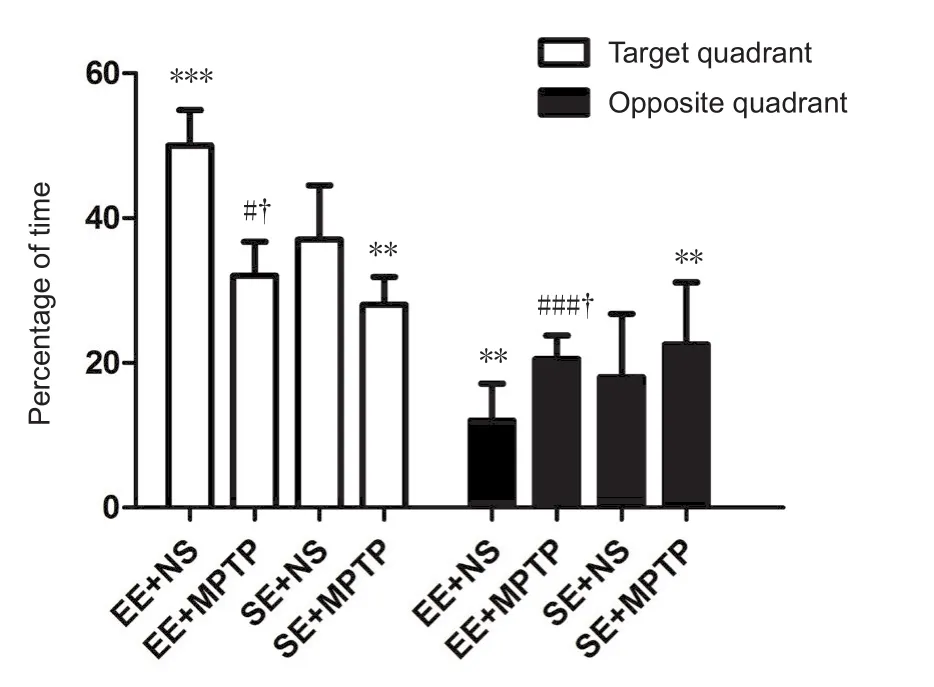
Figure 2 Effects of EE and MPTP on the swim time of SAMP8 mice in the spatial probe experiment of the Morris water maze task.
Compared with the SE + NS group, GAP-43-ir was significantly increased in the EE + NS group (P < 0.01). GAP-43-ir was significantly greater in the EE + MPTP group compared with the EE + NS group (P < 0.01) and was significantly greater in the SE + MPTP group compared with the SE + NS group (P <0.05). Compared with the SE + MPTP group, GAP-43-ir was greater in the EE + MPTP group (P < 0.01; Figure 4).
Effect of EE on GAP-43 protein levels in the substantia nigra of MPTP-treated mice
Compared with the SE + NS group, GAP-43 protein levels were significantly higher in the EE + NS group (P < 0.05).Compared with the EE + NS group, GAP-43 protein levels were significantly higher in the EE + MPTP group (P < 0.01).Compared with the SE + NS group, GAP-43 protein levels were significantly greater in the SE + MPTP group (P < 0.05).Compared with the SE + MPTP group, GAP43 protein levels were significantly increased in the EE + MPTP group (P< 0.01; Figure 5).
The man,7 who loved her dearly, thought to himself, Come! rather than let your wife die you shall fetch her some rampion, no matter the cost. 8 So at dusk he climbed over the wall into the witch s garden, and, hastily gathering2 a handful of rampion leaves, he returned with them to his wife. She made them into a salad, which tasted so good that her longing3 for the forbidden food was greater than ever.9 If she were to know any peace of mind, there was nothing for it but that her husband should climb over the garden wall again, and fetch her some more. So at dusk over he got, but when he reached the other side he drew back in terror, for there, standing4 before him, was the old witch.10
Discussion
With the number of PD patients increasing, an effective treatment method is desperately needed. Several studies have focused on new neuroprotective agents that may prevent dopaminergic cell loss (Reglódi et al., 2006, 2017; Song et al., 2012). Alternatively, there is evidence that exposure to EE improves neuroprotective effects (Young et al., 1999;Beauquis et al., 2010; Obiang et al., 2011), improves plasticity (Hosseiny et al., 2015; Mahati et al., 2016), reduces anxiety(Yuan et al., 2009b; Lach et al., 2016), ameliorates memory impairments (Yuan et al., 2013; Takuma et al., 2014; Wang et al., 2016; Bhagya et al., 2017), improves cognitive effects(Schreiber et al., 2014), and improves recuperation in animal models of PD (Requejo et al., 2018). EE refers to a living environment in which animals are kept in a large cage with running wheels, platforms, tunnels, and toys that provide increased opportunities for enhanced motor, cognitive and sensory stimulation (White et al., 2015; Huang et al., 2016).EE can dramatically induce behavioral and biochemical changes (van Praag et al., 2000; Kazlauckas et al., 2011), and influence the development and function of neural circuits(O’Connor et al., 2014). Physical exercise and sport can attenuate neuroin flammation (Spielman et al., 2016) and appear to be protective later in life (Lamotte et al., 2015; Shih et al., 2016). Appropriate environmental stimuli may help in the treatment of brain and behavioral dysfunction related to imbalances in the brain (Tomas et al., 2015). Although perinatal asphyxia and hypoxia can lead to permanent brain injuries (Placha et al., 2016; Rainaldi and Perlman, 2016), EE in early life can rescue dopaminergic cells after treatment of adult rats with 6-OHDA (Jungling et al., 2017).
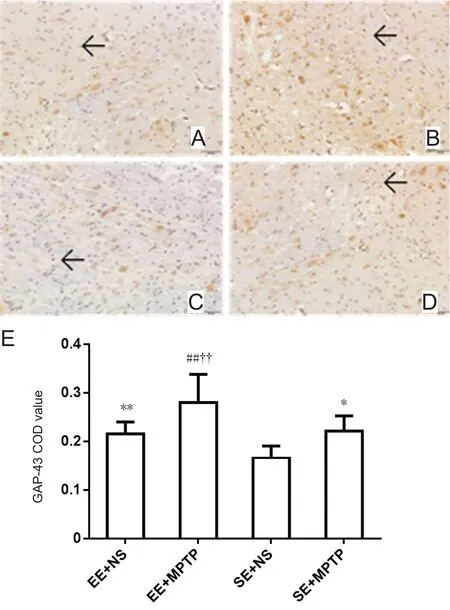
Figure 4 Effects of EE and MPTP on GAP-43-ir in the substantia nigra of SAMP8 mice.
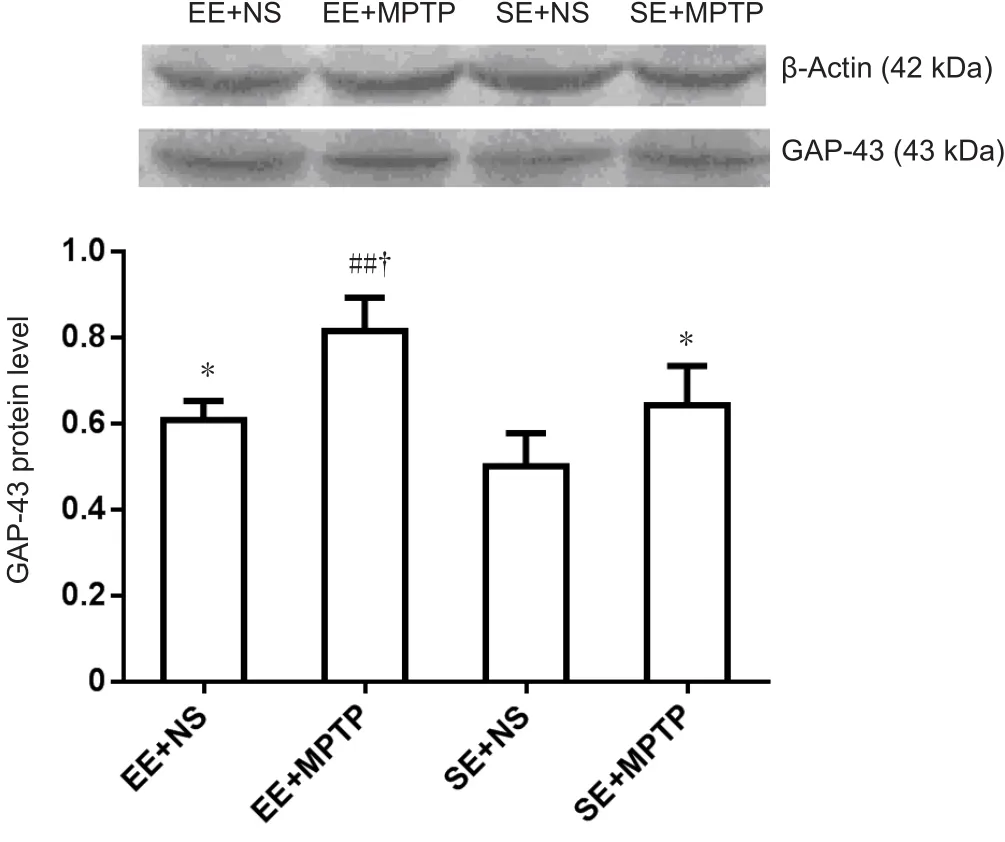
Figure 5 Effects of EE and MPTP on GAP-43 protein levels in the substantia nigra of SAMP8 mice.
Our previous study showed that EE can upregulate the mRNA and protein levels of brain-derived neurotrophic factor in the hippocampus (Yuan et al., 2012). The present study provides evidence that EE can improve cognitive ability and also increase the expression of GAP-43 in the substantia nigra of SAMP8 mice. This study assessed the cognitive ability of SAMP8 mice in different environments using the MWM test. Compared with the SE + NS group, escape latencies were shorter in the EE + NS group. Compared with the SE + MPTP group, escape latencies were shorter in the EE + MPTP group over four consecutive days. In the memory retention test, the percentage of time spent in the target quadrant was longer and the percentage of time spent in the opposite quadrant was shorter in the EE + NS group compared with the SE + NS group. Compared with the SE + MPTP group, the percentage of time spent in the target quadrant was longer and in the opposite quadrant was shorter in the EE + MPTP group. All these behavioral results showed that learning and retention performance was better in the EE group compared with the SE group, which indicated that EE might improve the cognitive abilities of mice in the water maze environment (Garthe et al., 2016). Compared with the SE group, greater space in the EE offered more opportunities for activity and more animals in a cage provided more social stimulation and mice also had enhanced opportunities for voluntary exercise on the running wheels. EE has a wide impact at the behavioral level and on molecular networks in the animal brain. Neuroplasticity changes are strongly associated with neurotrophic factors,neurotransmitters, neuronal patterns, brain network characteristics, and behavioral changes in cognition (Kotloski and Sutula, 2015).
The present study examined mRNA and protein levels of GAP-43, and GAP-43-ir. Compared with the SE + NS group,GAP-43 mRNA levels were increased in the EE + NS group.Compared with the SE + MPTP group, GAP-43 mRNA levels were higher in the EE + MPTP group. Compared with the SE+ NS group, GAP-43-ir was intense in the EE + NS group.Compared with the SE + MPTP group, corrected optical density values of GAP-43-ir were significantly increased in the EE + MPTP group. Furthermore, compared with the SE+ NS group, relative GAP43 protein levels were higher in the EE + NS group. Compared with the SE + MPTP group, GAP-43 protein levels were higher in the EE + MPTP group. All these results showed that EE upregulated the expression of GAP43 in the substantia nigra. Development of the brain and learning is accompanied by changes in the morphological structure of neurons and related molecular pathways. GAP-43 is a marker of neuronal elongation and synaptic formation that is highly expressed in the early stage of nervous system development (Rosskothen-Kuhl and Illing, 2014). Upregulation of GAP-43 and IGF-1 was found following treadmill running (Tsai et al., 2013). Voluntary exercise increased the levels of GAP-43 and enhanced recovery of cognitive function induced by traumatic injury (Gu et al., 2014). GAP-43 expression is strongest and most widespread during development (Jacobson et al., 1986), when the nervous system structure is subjected to extensive changes and rearrangements.The induction of GAP-43 was identified as an early event of neurogenesis (Chabot et al., 2013), and is strongly implicated in memory performance (Baumgartel and Mansuy, 2012).
In summary, EE improved learning and memory performance of SAMP8 mice and up-regulated GAP-43 mRNA and protein levels in the substantia nigra. This study provides a theoretical foundation of GAP-43 up-regulation being part of the mechanism for EE protection against MPTP-induced neuronal damage. However, the expression of GAP-43 in the striatum, hippocampus and cortex was not determined in the current study, and we will explore the expression of GAP-43 in different brain regions in future investigations.
Author contributions: ZYY and JY conceived and designed the study,wrote the paper, and were also responsible for data integration, analyses,and statistical analyses. XWM, YYW and MWW designed the study,and revised the paper. ZYY obtained the funding. All authors approved the final version of the paper.
Conflicts of interest: None declared.
Institutional review board statement: All animal experiments were approved by Animal Ethics Committee of Hebei Medical University in China.
Copyright license agreement: The Copyright License Agreement has been signed by all authors before publication.
Data sharing statement: Datasets analyzed during the current study are available from the corresponding author on reasonable request.
Plagiarism check: Checked twice by iThenticate.
Peer review: Externally peer reviewed.
Open access statement: This is an open access journal, and articles are distributed under the terms of the Creative Commons Attribution-Non-Commercial-ShareAlike 4.0 License, which allows others to remix, tweak,and build upon the work non-commercially, as long as appropriate credit is given and the new creations are licensed under the identical terms.
- 中国神经再生研究(英文版)的其它文章
- Restoration of an injured lower dorsal ascending reticular activating system in a patient with intraventricular hemorrhage
- Taurine protects against retinal and optic nerve damage induced by endothelin-1 in rats via antioxidant effects
- SIRT1 facilitates amyloid beta peptide degradation by upregulating lysosome number in primary astrocytes
- Cognitive deficits and Alzheimer-like neuropathological impairments during adolescence in a rat model of type 2 diabetes mellitus
- Achyranthes bidentata polypeptide protects dopaminergic neurons from apoptosis induced by rotenone and 6-hydroxydopamine
- Hyperoside protects the blood-brain barrier from neurotoxicity of amyloid beta 1–42