
渤海中国对虾生态容量变化研究*
2018-08-31 05:51单秀娟李忠义
渔业科学进展 2018年4期
林 群 单秀娟,2 王 俊 李忠义
渤海中国对虾生态容量变化研究*
林 群1单秀娟1,2王 俊1①李忠义1
(1. 农业农村部黄渤海渔业资源环境科学观测实验站 山东省渔业资源与生态环境重点实验室中国水产科学研究院黄海水产研究所 青岛 266071;2. 青岛海洋科学与技术试点国家实验室海洋渔业科学与食物产出过程功能实验室 青岛 266071)
基于1982年和2014~2015年渤海渔业资源与环境调查数据,采用Ecopath模型,分析了渤海生态系统的营养关系、结构及功能参数,评估了中国对虾()在渤海的生态容量变化。结果显示,渤海生态系统中底栖甲壳类、软体动物等功能群处于重要的营养位置,但中国对虾不是渤海生态系统的关键种,其生物量的增加对口虾蛄()、三疣梭子蟹()、多毛类、底栖甲壳类有负影响,花鲈()、虾虎鱼类等生物量的增加将对中国对虾产生负影响。渤海生态系统2个时期均处于发育的不稳定期,仍有较高的剩余生产量有待利用,2014~2015年渤海生态系统成熟度和稳定性较1982年有所降低,系统出现一定程度的退化。中国对虾1982年和2014~2015年在渤海的生态容量为0.810和0.702 t/km2;与当年依据调查数据评估的生物量相比较,中国对虾有较大的增殖潜力,当生物量增长至71.68倍和585倍时,仍不会超过生态容量。
渤海;中国对虾;营养关系;生态容量;Ecopath模型

1 材料与方法
1.1 模型原理与研究方法
渤海生态系统营养通道模型(Ecopath模型)利用建模软件Ecopath with Ecosim版本5.1和6.5 (Christensen, 2004)构建。Ecopath模型基于营养动力学原理,定义生态系统由一系列生态关联、能够基本覆盖生态系统能量流动过程的功能群(Group)组成,包含生态系统的所有生物组成部分,从碎屑和浮游植物到顶层捕食者(Polovina 1984; Christensen, 1992)。模型要求输入生物量()、生产量/生物量()、消耗量/生物量()和生态营养效率(EE) 4 个基本参数中的任意3个,以及食物组成矩阵和捕捞量参数。模型的调试是使生态系统每个功能群的输入和输出保持平衡,平衡满足的基本条件是:0<EE≤1。基本原理及调试方法具体参见林群等(2012)。
Ecopath模型利用混合营养分析程序(Mixed trophic impact,MTI)分析各功能群间的营养相互关系 (Leontief, 1951; Ulanowicz, 1990),基于MTI分析,提供了辨识生态系统关键种的方法(Power, 1996; Libralato, 2006),生态网络分析(Network analysis)功能可以评估表征生态系统结构和功能的生态指数、渔业捕捞对生态系统结构影响的相关指 数(Odum, 1969; Christensen, 2004)。
生态容量定义为特定时期、特定海域所能支持的,不会导致种类、种群以及生态系统结构和功能发生显著性改变的最大生物量,Ecopath模型为生态容量的研究提供了理论指导,基于海域的初级生产量基础,考虑了放流品种与食物竞争者、捕食者等间的相互作用,可评估生态容量,通过不断增加某放流品种的生物量(捕捞量也相应地成比例增加),观察系统中饵料生物等其他功能群的变化,当模型中任意其他功能群的EE>1时,模型将失去平衡而改变当前的状态,在模型即将不平衡前的放流品种生物量值即为生态容量(Jiang, 2005; Byron, 2011; 林群等, 2013)。
1.2 功能群的划分及数据来源
依据生物种类间的栖息地特征、生态学特征、简化食物网的研究策略(唐启升, 1999)以及评估中国对虾生态容量的研究目的,将渤海生态系统划分为19~20个功能群,包含重要渔业种群、中国对虾、中国对虾的食物竞争者和敌害生物,也包括有机碎屑、浮游植物、浮游动物、大型底栖动物、小型底栖动物等(表1),基本涵盖了渤海生态系统生物能量流动的组成成分。生物量、生产量和其他能量流动以湿质 量(t/km2)形式表示。
数据取自黄海水产研究所1982年2月、5月、8月、10月和2014年10月、2015年2月、5月、8月渤海渔业资源与环境调查,调查网具除网口周长有所不同,1982年为600目、2014~2015年为1740目,其他参数相同,网口高度6 m、宽22.6 m、网目63 mm、囊网网目20 mm,拖速大约3 n mile/h,所有调查数据均进行了标准化处理。对渔获物进行生物学测定和胃含物分析,鱼类、大型无脊椎动物的生物量通过扫海面积法(Gulland, 1965)计算得出,浮游植物生物量由叶绿素换算得出(Bundy, 2004; Lü, 2009),浮游动物生物量根据浮游动物大网采集数据,依据称重法(左涛等, 2003)计算得出。底栖动物利用箱式采泥器采样,大型底栖动物依据称重 法(张志南等, 1990; 张嵩等, 2017)测定生物量,小型底栖动物依据体积换算法(张青田等, 2011)测定生物量。碎屑量根据碎屑和初级生产碳的经验公式计 算(Christensen, 1993)。鱼类/值、/值利用经验公式计算得出(Pauly, 1980; Palomares, 1989),其他功能群种类的/、/值,以及其他未知参数等主要参考邻近水域模型(林群等, 2009、2012)。食性分析矩阵依据胃含物分析研究结果及已有的参考文献(邓景耀, 1988; 杨纪明, 2001; 张波等, 2004; Zhang, 2007)。捕捞量数据参考中国渔业统计年鉴(1982年, 2014年, 2015年),捕捞信息动态采集网络该研究水域调查的渔捞日志资料(http://www.eastfish.cn/index. aspx)以及渔民调研。EE值是较难获得的参数,由模型估算得出。
表1 渤海生态系统模型功能群的定义

Tab.1 Definition of functional groups in the Ecopath model of Bohai Sea ecosystem
2 结果与分析
2.1 模型参数质量与基本输出
Ecopath模型利用Pedigree指数分析数据来源和模型质量, 2个时期模型的Pedigree指数(Christensen, 2004)分别为0.495、0.507,均处于较合理范围,模型可信度较高。在系统的平衡过程中,微调输入的参数,同时进行了模型敏感度分析,2个模型的敏感度均在±20%范围内,通过鲁棒性检验。调试平衡的渤海Ecopath模型功能群的EE值均小于1,模型的输入参数和输出结果如表2所示,1982年和2014~2015年,中国对虾的营养级分别为3.11、3.22,生态营养效率分别为0.743、0.898。
表2 渤海Ecopath模型的基本输入参数和估计参数(黑体)

Tab.2 Basic input data and estimated parameters (bold) for Bohai Sea Ecopath model
注:“–”无对应数值
Note: “–” No corresponding value
2.2 营养相互关系与关键种分析
MTI分析是分析生态系统内部不同种群间营养关系的有效途径,渤海功能群间的营养相互关系利用MTI分析获得,中国对虾的食物竞争者、主要饵料生物和敌害生物等之间的营养关系在营养关系分析中可直观显示(图1)。1982年,中国对虾生物量的增加将对虾虎鱼类、花鲈产生正影响,影响值分别为0.0742、0.0544,对多毛类、软体动物和甲壳类产生负影响,影响值分别为–0.0207、–0.002、–0.0007;虾虎鱼类、花鲈、三疣梭子蟹、口虾蛄生物量的增加对中国对虾有负影响(–0.197、–0.119、–0.0145、–0.0052)。2014~2015年,中国对虾生物量的增加将对虾虎鱼类、其他底栖鱼类产生正影响,影响值分别为0.085、0.0042,对口虾蛄、多毛类、三疣梭子蟹和甲壳类产生较小负影响,分别为–0.002、–0.0012、–0.0005、–0.0005,虾虎鱼类生物量的增加对中国对虾有较大负影响(–0.673)。由于营养级联效应,中国对虾与其他的渔业捕捞功能群存在或多或少间接的影响。

图1 渤海生态系统功能群间的营养关系
渔业;其他功能群名见表2
Fishery; See Tab.2 for other functional groups
渤海生态系统各功能群按关键指数值排列顺序见对应图2。关键种对应有较高的总体效应、较高的关键指数(值接近或者大于0)的功能群。依据关键指数和总体效应值,1982年,浮游动物是渤海生态系统的重要种类,列入第1组;软体动物、浮游植物列入第2组;底栖甲壳类、其他底层鱼类列入第3组。2014~2015年,软体动物、底栖甲壳类列入第1组;蓝点马鲛、浮游动物、虾虎鱼类列入第2组;浮游植物列入第3组。这几个功能群在渤海海域生态系统中扮演着重要角色。
2.3 中国对虾的生态容量估算
在1982年和2014~2015年Ecopath模型基础上,分别评估了中国对虾的生态容量。1982年中国对虾现存生物量是0.0113 t/km2,当年5月中国对虾生物量是0.0017 t/km2。基于Ecopath模型估算生态容量的原理,大量放流中国对虾,当中国对虾生物量超过0.810 t/km2时,首先多毛类功能群EE≥1,随后底栖甲壳类功能群也将EE>1(表3),首先加大了对饵料生物的摄食压力,最终模型失去平衡。对比当前状态与放流中国对虾至0.810 t/km2时渤海生态系统的总体特征参数(表4),系统净生产量由882.76 t/km2·year降低为873.67 t/km2·year;总能量转换效率有所增加,由8.90%增加为9.80%,有效地提高了该海域基础生产力的利用率;渔业总捕捞效率有所提高,由0.0008提高到0.0015,渔获物的平均营养级由2.92增加到3.00;总初级生产量前后基本一致,无变化;系统其他能流与生态系统指数也基本差别不大,未改变系统其他组成的生物量与流动,未影响到水域系统的生态稳定性,由此确定渤海中国对虾的增殖生态容量为0.810 t/km2。

图2 渤海生态系统各功能群间的总体效应与关键指数
表3 估计中国对虾增殖生态容量(下划线)过程中模型的变动情况(1982年和2014~2015年)
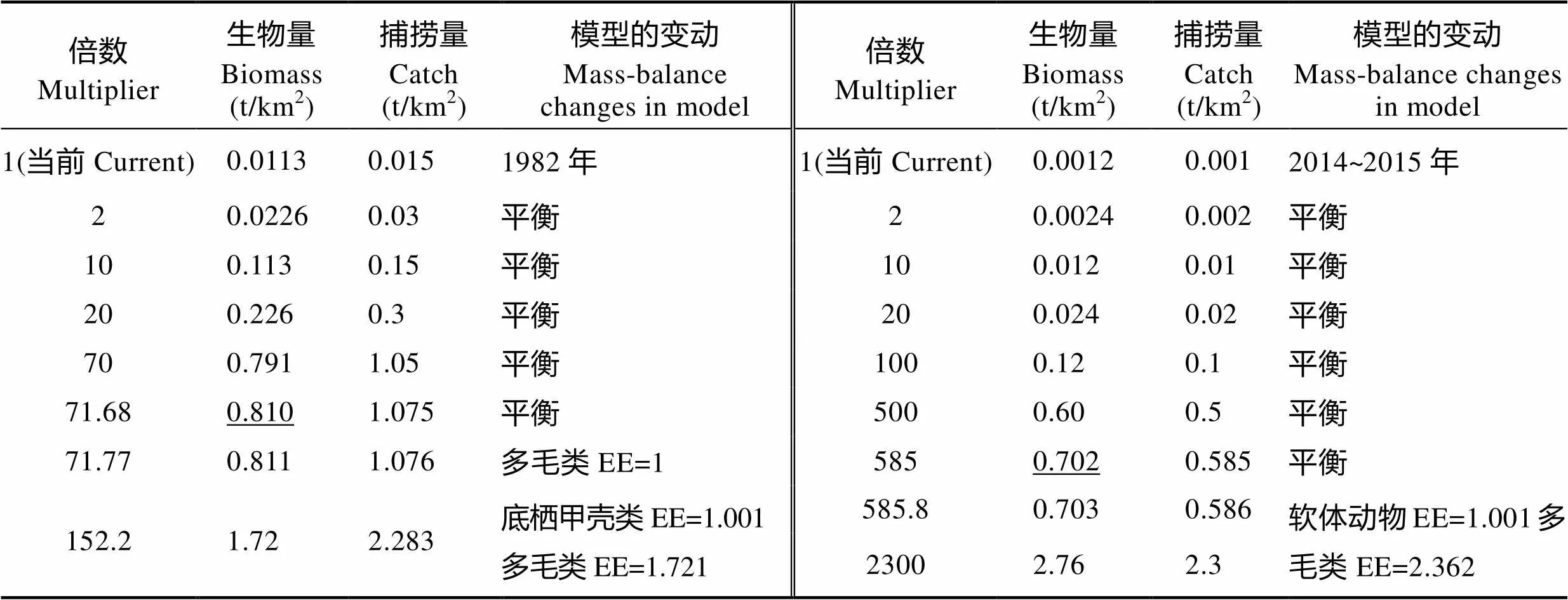
Tab.3 Changes in Ecopath model in estimating ecological carrying capacity of Chinese shrimp (underline) in the Bohai Sea ecosystem in 1982 and 2014~2015
表4 渤海生态系统的总体特征参数

Tab.4 General characteristic parameters for Bohai Sea ecosystem
注:V1,V2:当前的系统状态;V1¢,V2¢:放流中国对虾达到生态容量时的状态
Note: V1, V2: The current status of the system; V1¢, V2¢: The status at ecological carrying capacity of Chinese shrimp
2014~2015年中国对虾生物量为0.0012 t/km2,2015年5月未捕获到中国对虾,大量放流中国对虾,当中国对虾生物量超过0.702 t/km2时,首先多毛类功能群EE≥1, 随后软体动物功能群也将EE>1(表3),模型将失去平衡。对比当前状态与放流中国对虾生物量至0.702 t/km2时渤海生态系统的总体特征参数(表 4),系统净生产量由3922.88 t/km2·year降低为3914.88 t/km2·year;总能量转换效率有所增加,由5.10%增加为7.60%,有效地提高了该海域基础生产力的利用率;渔业总捕捞效率有所提高,由0.00018提高到0.00031,渔获物的平均营养级由2.68提高到2.90;总初级生产量前后基本一致,无变化;系统其他能流与生态系统指数也基本差别不大,未影响到水域系统的生态稳定性,由此确定渤海中国对虾的增殖生态容量为0.702 t/km2。
2.4 渤海生态系统结构和功能参数变化
基于生态网络分析,表4总结了渤海生态系统的一系列总体特征参数。系统总流量反映了系统规模大小,渤海生态系统的总体规模有所增加,2014~2015年系统总流量较1982年增加了1.44倍;流向碎屑总量也有所上升,2014~2015年的流向碎屑流量是1982年的3倍多,系统杂食系数稍微地降低,由0.15降为0.14,能量以碎屑形式在系统中的沉积增多。在成熟的生态系统中,系统没有多余的生产量再利用,总初级生产量与总呼吸量的比值(TPP/TR)接近1,总初级生产量与总生物量的比值(TPP/B)较低,1982年渤海生态系统TPP/TR为2.12,较2014~2015年的5.38低;1982年TPP/B为38.19,相比2014~2015年的99.83也较低,1982年和2014~2015年系统净生产量分别为882.76和3922.88 t/km2·year。渔获物的平均营养级(TLc)通过计算营养级获得捕捞种类在食物链中所处的相对位置,可以作为评价海洋生态系统渔业资源可持续利用的生态指标,也反映了鱼类、无脊椎动物等可捕种类种群结构的变化,捕捞渔获物的平均营养级从1982年的2.92下降到2014~2015年的2.68。
3 讨论
总体特征参数分析显示,2个时期渤海生态系统仍有较高的剩余生产量有待利用,系统均处于发育的不稳定期,易受外界干扰;2014~2015年与1982年相比,渤海生态系统浮游植物和碎屑的生物量明显增加,系统规模不断扩大,碎屑流在系统中的重要性逐渐降低,造成大部分剩余的能量在系统中沉积,一定程度地导致系统总体转换效率有所降低,系统的成熟度和稳定性较1982年有所降低。近些年海洋捕捞强度的不断加大、入海河流的陆源污染日趋严重以及栖息环境的变化等,是造成渤海生态系统成熟度和稳定性降低的主要原因。渤海生态系统捕捞渔获物的平均营养级由1982年的2.92下降到2014~2015年的2.68,主要原因是渔获物中高营养级生物比例下降, 而低营养级的小型生物比例有所上升,其他研究文献也证实了此结论。单秀娟等(2012)研究发现,目前渤海鱼类群落处于相对不稳定状态,渤海鱼类群落优势种由个体大、营养层次高的优质底层鱼类逐渐被个体小、营养层次低的小型中上层鱼类所替代。许思思 等(2014)认为渤海生态系统高营养级渔业生物群落逐渐衰退、低营养级渔业生物群落生物量相对增加是造成捕捞渔获物平均营养水平降低的根本原因,环境、上行饵料因素等是渤海捕捞渔获物平均营养级降低的次要因素。另外,渤海总捕捞产量从20世纪80年代中期开始一直保持上升趋势, 渔获物平均营养级的下降和捕捞强度的增加也紧密相关(林群等, 2016)。
中国对虾未成为渤海生态系统的关键种或者重要的功能群,中国对虾生物量的增加对食物竞争者(口虾蛄、三疣梭子蟹)、主要饵料生物(多毛类、软体动物、底栖甲壳类)有负影响,虾虎鱼类、花鲈幼鱼等均摄食少量中国对虾幼虾(唐启升等, 1997),花鲈、虾虎鱼类等生物量的增加将对中国对虾产生负影响。2014~2015年的软体动物、底栖甲壳类,1982年的浮游动物分别处于渤海生态系统的营养重要位置,关键作用发生了一定的转变;从营养关系的角度来看,中国对虾的增殖放流需兼顾生态系统间的营养平衡。以1982年和2014~2015年的Ecopath模型为基础,估算的中国对虾的增殖生态容量为0.810 t/km2和0.702 t /km2,与中国对虾现存生物量相比,渤海中国对虾有较大的增殖空间。受渤海生态系统退化的影响,2014~2015年中国对虾生态容量较1982年有所降低。历史上渤海秋汛中国对虾最高产量为4.1万t;按渤海海域面积77000 km2,依据现有生物量与捕捞量比例,对虾达到生态容量时,1982年和2014~2015年的捕捞产量分别为1.075 t/km2或82775 t、0.585 t/km2或45045 t,模型估算的2个时期中国对虾生态容量值对应的捕捞产量将超过历史的最高产量。上述方法估算的生态容量是从生态效益的角度考虑,是一个理论上限,依据渔业生产管理中采用的最大可持续产量(MSY)理论,最大生态容量值减半时,放流种群将获得较高生长率,因此指导中国对虾的增殖放流时需同时兼顾生态、经济、社会效益。
增殖放流的中国对虾仔虾主要摄食浮游植物,生长发育至成体后主要以底栖动物为食(邓景耀等, 1990)。当前,渤海生态系统浮游植物生物量与初级生产力水平较20世纪80年代有所增加,底栖生物较丰富,从饵料角度考虑,中国对虾饵料较充足,放流期间饵料生物等级较高,有利于放流个体成活率的提高;敌害生物对中国对虾的危害主要在近岸,水深5 m以下水域受敌害生物的影响较小,敌害生物资源密度相对较低(单秀娟等, 2012; 吴强等, 2016)。但调查显示,2015年春季基本捕捞不到中国对虾,中国对虾群落结构发生较大变化。邓景耀等(2001)认为,恢复和增加渤海对虾补充量是重建渤海对虾渔业的有效途径,20世纪80年代对虾育苗养殖业的兴起造成对虾亲虾的严重不足、对虾栖息环境的破坏、对虾遗传多样性的下降是造成中国对虾补充量下降、锐减的直接原因。因此,增殖放流的数量控制至关重要,适度增殖同时保护幼虾放流的水域环境、提高遗传多样性,进而提高中国对虾的补充量,实现增殖放流的生态型放流目标。
4 结论
综上所述,中国对虾未成为渤海生态系统的关键种或者重要的功能群,以1982年和2014~2015年的Ecopath模型为基础,估算的中国对虾的增殖生态容量为0.810 t/km2和0.702 t/km2,与通过调查评估的中国对虾生物量相比,渤海中国对虾有较大的增殖空间。2014~2015年渤海生态系统成熟度和稳定性较1982年有所降低,系统出现一定程度的退化,2014~ 2015年中国对虾生态容量较1982年有所降低。建议渤海对虾增殖放流时,采用中国对虾生物量至最大生态容量值减半,来指导增殖生产。
Aprahamian MW, Martin SK, Mcginnitw P,. Restocking of salmonids opportunities and limitations. Fisheries Research, 2003, 62(2): 211–227
Blankenship HL, Leber KM. A responsible approach to marine stock enhancement. American Fisheries Society Symposium, 1995, 15: 167–175
Bundy A. Mass balance models of the eastern Scotian Shelf before and after the cod collapse and other ecosystem changes. Canadian Technical Report of Fisheries and Aquatic Sciences, 2004, 2520: xii–193
Byron C, Link J, Costa-Pierce B,. Calculating ecological carrying capacity of shellfish aquaculture using mass- balance modeling: Narragansett Bay, Rhode Island. Ecological Modelling, 2011, 222(10): 1743–1755
Bell JD, Rothlisberg PC, Munro JL,. Restocking and stock enhancement of marine invertebrate fisheries. Advances in Marine Biology, 2005, 49: xi–374
Blaxter JHS. The enhancement of cod stocks. Fish & Fisheries, 2000, 1(2): 173–205
Christensen V, Pauly D. ECOPATH II-a software for balancing steady-state ecosystem models and calculating network characteristics. Ecological Modelling, 1992, 61(3–4): 169– 185
Christensen V, Walters CJ, Pauly D. Ecopath with Ecosim: A user’s guide. Fisheries Centre of University of British Columbia, Vancouver, Canada, 2004. 154 p
Deng JY, Meng TX, Ren SM. Food web of fishes in Bohai Sea. Marine Fisheries Research, 1988(9): 151–172 [邓景耀, 孟田湘, 任胜民. 渤海鱼类的食物关系. 海洋水产研究, 1988(9): 151–172]
Deng JY, Ye CC, Liu YC. The Chinese shrimp () and its resources management in Bohai Sea and the Yellow Sea. Beijing: Ocean Press, 1990 [邓景耀, 叶昌臣, 刘永昌. 渤黄海的对虾及其资源管理. 北京: 海洋出版社, 1990]
Deng JY, Zhuang ZM. The cause of recruitment variation of Penaeus chinensis in the Bohai Sea. Journal of Fishery Science of China, 2000, 7(4): 125–128 [邓景耀, 庄志猛. 渤海对虾补充量变动原因的分析及对策研究. 中国水产科学, 2000, 7(4): 125–128]
Guo XP, Jin XS, Dai FQ. Growth variations of small yellow croaker () in the Bohai Sea. Journal of Fishery Sciences of China, 2006, 13(2): 243−249 [郭旭鹏, 金显仕, 戴芳群. 渤海小黄鱼生长特征的变化. 中国水产科学, 2006, 13(2): 243−249]
Gulland JA. Manual of methods for fish stock assessment. Part 1. Fish population analysis. FAO Fisheries Technical Paper, 1965, (40): 72
Jiang W, Gibbs MT. Predicting the carrying capacity of bivalve shell- fish culture using a steady, linear food web model. Aquaculture, 2005, 244(1–4): 171–185
Jin XS. The dynamics of major fishery resources in the Bohai Sea. Journal of Fishery Sciences of China, 2001, 7(4): 22–26 [金显仕. 渤海主要渔业生物资源变动的研究. 中国水产科学, 2001, 7(4): 22–26]
Jin XS. Long-term changes in fish community structure in the Bohai Sea, China. Estuarine, Coastal and Shelf Science, 2004, 59: 163–171
Jin XS, Qiu SY, Liu XZ,. The foundation and prospect of stock enhancement of fishery resources in the Bohai Sea and Yellow Sea. Beijing: Science Press, 2014: 395 [金显仕, 邱盛尧, 柳学周, 等. 黄渤海渔业资源增殖基础与前景. 北京: 科学出版社, 2014: 395]
Leontief WW. The structure of the U. S. economy. New York: Oxford University Press, 1951
Libralato S, Christensen V, Pauly D. A method for identifying keystone species in food web models. Ecological Modelling, 2006, 195(3): 153–171
Lin Q, Jin XS, Zhang B,. Comparative study on the changes of the Bohai Sea ecosystem structure based on Ecopath model between ten years. Acta Ecologica Sinica, 2009, 29(7): 3613–3620 [林群, 金显仕, 张波, 等. 基于营养通道模型的渤海生态系统结构十年变化比较研究. 生态学报, 2009, 29(7): 3613–3620]
Lin Q. Studies on the ecosystem energy transfer and function in the typical waters of Yellow and Bohai Sea. Doctoral Dissertation of Ocean University of China, 2012, 1–127 [林群. 黄渤海典型水域生态系统能量传递与功能研究. 中国海洋大学博士研究生学位论文, 2012, 1–127]
Lin Q, Li XS, Li ZY,. Ecological carrying capacity of Chinese shrimp stock enhancement in Laizhou Bay of East China based on Ecopath model. Chinese Journal of Applied Ecology, 2013, 24(4): 1131–1140 [林群, 李显森, 李忠义, 等.基于Ecopath 模型的莱州湾中国对虾增殖生态容量.应用生态学报, 2013, 24(4): 1131–1140]
Lin Q, Wang J, Yuan W,. Effects of fishing and environmental change on the ecosystem of the Bohai Sea. Journal of Fishery Sciences of China, 2016, 23(3): 619–629 [林群, 王俊, 袁伟, 等. 捕捞和环境变化对渤海生态系统的影响. 中国水产科学, 2016, 23(3): 619–629]
Lorenzen K, Leber KM, Blankenship HL. Responsible approach to marine stock enhancement: An update. Reviews in Fisheries Science, 2010, 18(2): 189–210
Lü SG, Wang XC, Han BP. A field study on the conversion ratio of phytoplankton biomass carbon to chlorophyll-a in Jiaozhou Bay, China. Chinese Journal of Oceanology and Limnology, 2009, 27(4): 793–805
Mcdowell N. Stream of escaped farm fish raises fears for wild salmon. Nature, 2002, 416(6881): 571
Odum EP. The strategy of ecosystem development. Science, 1969, 164: 262–270
Palomares MLD, Pauly D. A multiple regression model for predicting the food consumption of marine fish populations. Marine & Freshwater Resources, 1989, 40(3): 259–273
Pauly D. On the interrelationships between natural mortality, growth parameters, and mean environmental temperature in 175 fish stocks. ICES Journal of Marine Science, 1980, 39(2): 175–192
Polovina JJ. Model of a coral reef ecosystem I: The ECOPATH model and its application to French Frigate Schoals. Coral Reefs, 1984, 3(12): 1–11
Power ME, Tilman D, Estes JA,. Challenges in the quest for keystones. Bioscience, 1996, 46 (8): 609–620
Shan XJ, Jin XS. Population dynamics of fish species in a marine ecosystem: A case study in the Bohai Sea, China. Marine and Coastal Fisheries: Dynamics, Management, and Ecosystem Science, 2016, 8: 100–117
Shan XJ, Jin XS, Li ZY,. Fish community structure and stock dynamics of main releasing fish species in the Bohai Sea. Progress in Fishery Sciences, 2012, 33(6): 1–9 [单秀娟, 金显仕, 李忠义, 等. 渤海鱼类群落结构及其主要增殖放流鱼类的资源量变化. 渔业科学进展, 2012, 33(6): 1–9
Tang QS. Strategies of research on marine food web and trophodynamics between high trophic levels. Marine Fisheries Research, 1999, 20(2): 1–11 [唐启升. 海洋食物网与高营养层次营养动力学研究策略. 海洋水产研究, 1999, 20(2): 1–11]
Tang QS, Jin XS, Wang J,. Decadal-scale variations of ecosystem productivity and control mechanisms in the Bohai Sea. Fisheries Oceanography, 2003, 12(4–5): 223–233
Tang QS, Wei S, Jiang WM. Predator species of fishery resources enhancement and their predation on enhancement species in Laizhou Bay of Bohai Sea. Chinese Journal of Applied Ecology, 1997, 8(2): 199–206 [唐启升, 韦晟, 姜卫民. 渤海菜州湾渔业资源增殖的敌害生物及其对增殖种类的危害. 应用生态学报, 1997, 8(2): 199–206]
Ulanowicz RE, Norden JS. Symmetrical overhead in flow networks. International Journal of Systems Science, 1990, 21(2): 429–437
Wu Q, Jin XS, Luan QS,. Analysis on the reproduction ofandbased on their food sources and predators in the Laizhou Bay of China. Progress in Fishery Science, 2016, 37(2): 1–9 [吴强, 金显仕, 栾青杉, 等. 基于饵料及敌害生物的莱州湾中国对虾()与三疣梭子蟹()增殖基础分析. 渔业科学进展, 2016, 37(2): 1–9
Xu SS, Song JM, Li XG,. Variation characteristics of catch structure in the Bohai Sea and its influencing factors. Journal of Natural Resources, 2014, 29(3): 500–506 [许思思, 宋金明, 李学刚, 等. 渤海渔获物资源结构的变化特征及其影响因素分析. 自然资源学报, 2014, 29(3): 500– 506]
Yang JM. A study on food and trophic levels of Bohai Sea invertebrates. Modern Fisheries Information, 2001, 16(9): 8–16 [杨纪明. 渤海无脊椎动物的食性研究. 现代渔业信息, 2001, 16(9): 8–16]
Zhang B, Tang QS. Study on trophic level of important resources species at high trophic levels in the Bohai Sea, Yellow Sea and East China Sea. Advances in Marine Science, 2004, 22(4): 393–404[张波, 唐启升. 渤、黄、东海高营养层次重要生物资源种类的营养级研究. 海洋科学进展, 2004, 22(4): 393–404]
Zhang B, Tang QS, Jin XS. Decadal-scale variations of trophic levels at high trophic levels in the Yellow Sea and the Bohai Sea ecosystem. Journal of Marine Systems, 2007, 67: 304–311
Zhang ZN, Tu LH, Yu ZS. Preliminary study on themacrofauna in the Huanghe River estuary and its adjacent waters (I) the biomass. Journal of Ocean University of Qingdao, 1990, 20(1): 37–45 [张志南, 图立红, 于子山. 黄河口及其邻近海域大型底栖动物的初步研究(一)生物量. 青岛海洋大学学报自然科学版, 1990, 20(1): 37–45]
Zhang S, Zhang CL, Xu BD,. Ecological status assessment based on macrobenthic community characteristics in Yellow River estuary and its adjacent waters. Periodical of Ocean University of China, 2017, 47(5): 65–71 [张嵩, 张崇良, 徐宾铎, 等. 基于大型底栖动物群落特征的黄河口及邻近水域健康度评价. 中国海洋大学学报自然科学版, 2017, 47(5): 65–71]
Zhang QT, Hu GK. Review and thinking about the estimation method of meiofaunal biomass. Marine Science Bulletin, 2011, 30(3): 357–360 [张青田, 胡桂坤. 小型底栖动物生物量估算方法回顾与思考. 海洋通报, 2011, 30(3): 357–360]
Zuo T, Wang R. Measuring methods of marine zooplankton biomass. Chinese Journal of Ecology, 2003, 22(3): 79–83 [左涛, 王荣. 海洋浮游动物生物量测定方法概述. 生态学杂志, 2003, 22(3): 79–83]
(编辑 江润林)
Changes in Chinese Shrimp () Carrying Capacity of the Bohai Sea
LIN Qun1, SHAN Xiujuan1,2, WANG Jun1①, LI Zhongyi1
(1. The Yellow & Bohai Seas Scientific Observation and Experiment Station for Fishery Resources and Environment, Ministry of Agriculture and Rural Affairs, Shandong Provincial Key Laboratory of Fishery Resources and Eco-Environment, Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Qingdao 266071; 2. Laboratory for Marine Fisheries Science and Food Production Processes, Pilot National Laboratory for Marine Science and Technology (Qingdao), Qingdao 266071)
In this study, data from surveys of fishery resources and environments collected from the Bohai Sea ware used to construct an Ecopath model of 1982 and 2014~2015, respectively. Based on the models, we analyzed the trophic impact relationship, structure, and functional parameters of the Bohai Sea ecosystem. Furthermore, we calculated the ecological carrying capacity for Chinese shrimp in the Bohai Sea and compared its changes between the two periods. Trophic impact and keystoness index results showed that the Chinese shrimp was not a keystone species or an important functional group of this ecosystem. Benthic crustaceans, mollusks, and other functional groups occupied important trophic positions in the Bohai Sea ecosystem. The increase in Chinese shrimp biomass will have a negative impact on,, other benthic crustaceans, and polychaetes. The increase in the biomass of functional groups such asand gobies may have a negative impact on Chinese shrimp. Network analysis showed that this ecosystem was at an unstable developmental stage during these two periods, and the Bohai Sea ecosystem had higher surplus production. A comparison of the 1982 and 2014~2015 models showed that the maturity and stability of the ecosystem decreased, resulting from its degeneration. The carrying capacity of Chinese shrimp was found to be 0.810 and 0.702 t/km2in Bohai Sea in 1982 and 2014~2015, respectively. Compared with its current biomass, Chinese shrimp had great potential for continuous enhancement, e.g., by releasing larvae in the Bohai Sea, and the ecological carrying capacity was not exceeded even when the biomass of the Chinese shrimp was increased by 71.68 and 585 fold in 1982 and 2014~2015, respectively. This study provides information for rational implementation of stock enhancement and promotion of sustainable development of fisheries resources in this sea area.
Bohai Sea;; Trophic impact relationship; Ecological carrying capacity; Ecopath model
WANG Jun, E-mail: wangjun@ysfri.ac.cn
10.19663/j.issn2095-9869.20170908001
S 931.1
A
2095-9869(2018)04-0019-11
* 国家重点基础研究发展计划项目(2015CB453303)、公益性行业(农业)科研专项经费(201303050)、中海油公益基金项目渤海对虾容量、山东省泰山学者建设工程专项和青岛海洋科学与技术国家实验室“鳌山人才”计划共同资助[This work was supported by the National Basic Research Program of China (2015CB453303), the Special Fund for Agro-Scientific Research in the Public Interest (201303050), China National Offshore Oil Corporation Public Welfare Fund Project-Fleshy Prawn Capacity of Bohai Sea, Ministry of Agriculture and the Special Fund of the Taishan Scholar Project and Talent Plan of Qingdao National Laboratory for Marine Science and Technology]. 林 群, E-mail: linqun@ysfri.ac.cn
王 俊,研究员,E-mail: wangjun@ysfri.ac.cn
2017-09-08,
2018-03-22
林群, 单秀娟, 王俊, 李忠义. 渤海中国对虾生态容量变化研究. 渔业科学进展, 2018, 39(4): 19–29
Lin Q, Shan XJ, Wang J, Li ZY. Changes in Chinese shrimp () carrying capacity of the Bohai Sea. Progress in Fishery Sciences, 2018, 39(4): 19–29
猜你喜欢
声屏世界(2022年17期)2022-12-18
中国交通信息化(2022年7期)2022-10-27
当代水产(2022年8期)2022-09-20
当代水产(2022年5期)2022-06-05
当代水产(2021年8期)2021-11-04
当代水产(2021年4期)2021-07-20
今日农业(2020年19期)2020-12-14
水上消防(2020年3期)2020-07-25
科技进步与对策(2020年9期)2020-05-28
安徽农学通报(2020年7期)2020-05-26
