
干热河谷冲沟沟岸葛藤不同覆被状况及土壤性质差异分析
2018-08-29 11:08熊东红张宝军
生态学报 2018年14期
校 亮,熊东红,张宝军,张 素,吴 汉,杨 丹
1 中国科学院山地灾害与地表过程重点实验室,成都 610041 2 中国科学院水利部成都山地灾害与环境研究所,成都 610041 3 中国科学院大学资源与环境学院,北京 100049
植被通过根系与土壤连成统一整体,并通过代谢活动影响土壤性质及其演化[1],对土壤性质及其质量影响显著[2-3]。植被恢复主要通过物理作用[4](如覆被遮荫、截流入渗、根系穿插)和化学作用[5](如残体分解与养分归还、根系分泌物与土壤胶结)等方式直接或间接影响着土壤水分物理环境及化学性质,并进一步影响土壤生物活性[6]。
金沙江干热河谷是我国土壤生态环境脆弱区,也是植被恢复困难区[7]。1950 s以来,为防治水土流失、改善生态环境,先后开展了一系列植被恢复的理论研究和实践探索,涵盖植被恢复的限制性因子[8]、适宜树种的筛选与引种[9]、植被恢复的技术方法[10]、有效性评估等多个方面[11-12]。如Tang等[13]指出,新银合欢(Leucaenaleucocephala)在干热河谷具有较好的固碳潜力;Forrester等[14]研究表明,含羞草科植物能有效固定大气氮素以满足植物生长,进而促进土壤改良;唐国勇等[15]发现植被类型和恢复年限决定土壤改良进程。但是,已有研究多集中在流域或坡地尺度,针对裸露、陡立的冲沟沟岸的植被恢复及其对土壤性质的影响研究却少有报道,罗君等针对元谋干热河谷冲沟区植被对微地形的响应研究中,其指出燥红土区活跃性冲沟坡面植被生长的临界坡度为小于65°,植被盖度和种类的优势方向同为东北向[16]。
沟岸是冲沟发育过程中因流水下切侵蚀和侧壁崩塌发育形成的,由于特殊的微地形条件,降雨过程中其水分入渗及保存困难,并长期受阳光直射,水热矛盾极为突出;同时,沟岸土壤退化严重、土层浅薄、土壤贫瘠;因而植被恢复极端困难,长期以来一直未能找到有效的植被恢复措施。而探寻适宜沟岸的植被恢复措施、并深入开展植被生长对沟岸土壤性质影响的研究,对于深刻认识土壤-植被相互作用机理、指导冲沟活跃地带的植被恢复实践及控制水土流失具有重要意义。
本文以元谋干热河谷区冲沟发育的沟岸土体为研究对象,基于野外调查,选取藤本植物葛藤(Kudzu)覆被为沟岸植被恢复措施,结合室内实验,定量分析葛藤不同覆被状况下沟岸土壤性质的变化特征,探讨葛藤生长对沟岸土壤性质的影响,旨在为干热河谷冲沟系统生境改良与植被恢复提供有益参考。
1 材料与方法
1.1 研究区概况
研究区位于元谋干热河谷区(101°35′—102°26′E,25°23′—26°06′N),该区属南亚热带季风气候,具有“炎热干燥、降雨集中、干湿季分明”的气候特征[17];区内地带性土壤为燥红土;自然植被为稀树灌木草丛[18]。该区冲沟发育,沟壑密度为3.0—5.0 km/km2,最高达7.4 km/km2,因下切侵蚀和侧壁崩塌,沟谷两侧常发育有陡立沟岸[19]。
1.2 样地选取
本研究所指的冲沟沟岸,区分于沟头以上及其临近范围受集水区径流冲刷影响的区域,特指在地形平坦开阔的沟床两侧的陡立地带,即从沟床侧壁底部向上延伸至沟缘的区域。其地形特征在于地形坡度十分陡峭,不利于植被生长,地表常常裸露;由于特殊的地形和植被状况,使得降雨过程中水分入渗及保存困难,并长期受阳光直射的影响,水热矛盾极为突出,导致土壤退化严重,土层浅薄、土壤贫瘠,植被生长更加困难。
2016年3月,在元谋沟蚀崩塌观测研究站(101°83′E,25°85′N)附近选取了一葛藤生长的原位沟岸。葛藤属多年生藤本植物,2006年种植在沟底区域,攀沿生长以布满沟岸,因在沟底葛藤的种植密度和管护措施等的差异,不同地块覆被状况各异,因处旱季时段,出现了大量枯藤和极少鲜藤共存的景象。沟岸坡向30°、坡度67.5°、坡面长5.9 m、岸宽47 m,燥红土层(上层3.9 m)、砂积层(下层2 m)成层分布。
研究采用空间替代时间的方法,探讨葛藤不同生长阶段土壤性质变化特征及其差异成因。随葛藤生活史的进行,试验沟岸会经历裸露→鲜藤覆被→枯藤覆被→枯藤逐年累积等阶段。
基于葛藤自然生长状况,在沟岸沿坡面长2 m以下的燥红土层选取试验地块(规格1.8 m×1.8 m),以规避沟缘以上台面区域水分入渗/蒸散对深层土壤水分的影响,尽可能地保证沟岸土壤水分散失只取决于侧壁蒸发。其中选取裸露地块作对照CK,鲜藤覆被地块设置1种处理T1,枯藤覆被地块设置3种处理(T2、T3、T4),葛藤覆被状况以单位面积生物量计(图1)。

图1 试验区原位小区处理图示Fig.1 Sketch of experimental field plotsCK:裸露沟岸, the bare gully bank; T1: 鲜藤覆被沟岸, the gully bank under fresh coverage of Kudzu; T2—T4: 枯藤覆被沟岸, the gully bank under withered coverage of Kudzu
1.3 葛藤生物量测定
采集各地块葛藤覆被物,各处理取4个重复样(S形采样)。将自制方形环刀(20 cm×20 cm×20 cm)垂直轻扣在覆被物上,紧贴环刀外壁刈割覆被物,并收集环刀内所有覆被物。烘干法(85℃)测定其生物量,算数平均后换算求得葛藤单位面积生物量,作为葛藤覆被状况指标(表1)。

表1 试验处理设置
CK:裸露沟岸, the bare gully bank; T1: 鲜藤覆被沟岸, the gully bank under fresh coverage ofKudzu; T2—T4: 枯藤覆被沟岸, the gully bank under withered coverage ofKudzu
1.4 土壤样品采集与分析
2016年3月进行土样采集。用环刀法取0—10 cm表层土壤,分别于2016年4月下旬、5月上旬、中旬采集,每次按S形取5个点的混合样500 g左右,放入聚乙烯袋中密封带回,随即置于0℃冰箱中冷藏保存,以备室内分析测定各种指标,各土壤性质分析指标及测定方法如表2所示[20]。
其中,试验土壤微生物量测定采用PLFA法,测定指标包括单位质量土壤总PLFA量,单位质量土壤中的细菌、真菌、放线菌、原生动物的相对含量百分比,前者是表征土壤微生物量的重要指标,后者反映了土壤微生物群落结构及其活性[21]。PLFA测定的是表征微生物群落信息的磷脂脂肪酸甲酯,前处理将磷脂脂肪酸提取出来,甲酯化后通过GC分析。数据库匹配信息后,给出检测到的甲酯的名称及其含量百分比。上机前加入定量C19甲酯内标,通过内标计算土壤中每种甲酯的质量含量,不同的甲酯表征不同的微生物的种类及其含量。表征土壤微生物的PLFA如表3所示[21]。

表2 土壤性质分析指标及测定方法

表3 表征土壤微生物的PLFAs
1.5 数据统计与分析
采用EXCEL 2013和SPSS 13.0进行相关数据的整合处理与统计分析,所有数据通过正态分布检验和方差一致性检验,用一维方差分析(one-way ANOVA)检验葛藤不同覆被状况下各土壤性质参数变化的显著性(P<0.05)。
2 结果与分析
2.1 葛藤覆被对沟岸土壤物理性状的影响
土壤是由固、液、气及多种活的有机体组成的一个十分复杂的无机-有机复合系统,它作为植物生长的基质,不仅要供给植物生长所需要的水分,还要向根系提供养分和氧气,这些不同的功能主要取决于土壤固相的组成和性质,即土壤物理性状[22]。
植被生长对土壤物理性状影响显著,其主要通过截流入渗、根系穿插等方式改变土壤物理性状,尤其对土壤水分状况及孔隙结构有较大影响[23-24]。表4反映了葛藤不同覆被状况下沟岸表层0—10 cm土壤物理性状(土壤含水率、容重、毛管孔隙度、非毛管孔隙度)的变化差异。

表4 葛藤不同覆被状况下沟岸土壤物理性状差异
表内±后跟数据为标准差;同列数据中标有相同的字母表示其统计差异不显著(P> 0.05)
土壤水分是沟岸植被恢复与重建的重要影响因子,水分供给不足将限制植被生长甚至威胁其生存,它直接反映了土壤供给植物生存、生长的基本能力[25]。研究发现,葛藤不同覆被状况下沟岸土壤含水率变化差异明显,相比裸露沟岸而言(VCK=11.05%),鲜藤覆被沟岸土壤含水率明显降低,仅为7.63%;枯藤覆被下沟岸土壤含水率持续增加,且枯藤覆被量越多,其下土壤含水率越高;具体地,T2—T4处理沟岸土壤含水率依次为13.30%、16.49%、18.90%。方差分析表明,鲜、枯藤覆被沟岸土壤含水率变化差异显著。上述研究结果表明沟岸土壤水分状况受葛藤覆被类型(鲜/枯)及覆被物生物量共同影响,枯藤覆被有助于保持其下沟岸土壤水分,鲜藤覆被下沟岸土壤水分则明显减小。
土壤容重综合反映孔隙状况、松紧度和肥力等,对土壤透气状况、入渗性能、持水能力等均有很大影响[26]。研究发现,相比裸露沟岸而言,鲜藤覆被沟岸土壤容重相对减小,T1处理较CK对照土壤容重减小了3.92%;在枯藤覆被沟岸,随枯藤覆被量的增加,沟岸土壤容重依次降低(T2、T3、T4土壤容重依次为1.59、1.54、1.50 g/cm3),但均高于裸露沟岸(1.45 g/cm3)。方差分析表明,较裸露沟岸,鲜、枯藤覆被沟岸土壤容重变化差异显著;在枯藤覆被沟岸,T2—T4处理沟岸土壤容重存在差异但差异不显著。上述研究结果表明沟岸土壤容重同样受葛藤覆被类型(鲜/枯)及覆被物生物量共同影响,鲜藤覆被有助于降低其下沟岸土壤容重,枯藤覆被下沟岸土壤容重反而增加。
土壤孔隙直接影响土壤通气透水性能、水分保持与运移、根系穿插难易等。其中,土壤毛管孔隙度反映了植被吸持水分用于维持自身生长发育的能力,非毛管孔隙度反映了植被滞留水分发挥涵养水源的能力等[27]。研究发现,相比裸露沟岸而言,鲜、枯藤覆被沟岸土壤毛管孔隙度均有所增加,尤其以鲜藤覆被沟岸土壤毛管孔隙度增加最为明显。具体地,T1—T4处理较CK对照毛管孔隙度分别增加了18.85%、8.18%、8.91%、10.17%。在枯藤覆被沟岸,T2—T4处理毛管孔隙度和非毛管孔隙度均随枯藤覆被量的增加而增加。T2—T4处理土壤毛管孔隙度依次为40.48%、40.75%、41.22%,非毛管孔隙度依次为1.26%、1.32%、2.96%。方差分析表明,较裸露沟岸,鲜、枯藤覆被沟岸土壤毛管孔隙度、非毛管孔隙度变化差异显著;在枯藤覆被沟岸,T4较T2、T3处理沟岸土壤非毛管孔隙度变化差异显著。上述研究结果表明沟岸土壤孔隙状况依旧受葛藤覆被类型(鲜/枯)及覆被物生物量共同影响。鲜、枯藤覆被均有助于增加其下沟岸土壤毛管孔隙度,但均不利于土壤非毛管孔隙度的提升。
2.2 葛藤覆被对沟岸土壤化学性质的影响
植被恢复显著影响土壤化学性质,其主要通过提供生物残体、产生根系分泌物、活化土壤微生物等方式来影响土壤酸碱度、土壤胶体性质及土壤养分状况等[28],对植物生长下的土壤化学特性进行分析,是改良土壤、提高肥力、补充作物所需营养元素的前提和基础。表5反映了葛藤不同覆被状况下沟岸表层0—10 cm土壤化学性质(全盐含量、有机质、铵态氮、硝态氮、有效磷、速效钾)的变化差异。

表5 葛藤不同覆被状况下沟岸土壤化学性质差异
表内±后跟数据为标准差;同列数据中标有相同的字母表示其统计差异不显著(P> 0.05)
研究发现,相比裸露沟岸而言,鲜、枯藤覆被沟岸土壤全盐含量均呈现整体增加的规律。具体表现为,相比裸露沟岸而言,鲜(T1)、枯藤覆被(T2—T4)沟岸土壤全盐含量分别增加了199.45%、66.08%、198.20%、204.16%。方差分析表明,较裸露沟岸,鲜、枯藤覆被沟岸土壤全盐含量变化差异显著(T2处理除外);在枯藤覆被沟岸,T3、T4处理间沟岸土壤全盐含量变化无差异,但T3和T4处理较T2处理沟岸土壤全盐含量变化差异显著。上述变化特征表明鲜、枯藤覆被均加重了其下沟岸土壤受盐分的胁迫作用。
研究发现,相比裸露沟岸而言,鲜藤覆被沟岸土壤有机质含量相对降低,而枯藤覆被沟岸土壤有机质含量则明显增加,且枯藤覆被量越多,增幅越显著。具体地,较裸露沟岸,鲜藤覆被沟岸土壤有机质含量降低了25.93%,而在枯藤覆被沟岸,T2—T4处理土壤有机质含量分别提升了41.20%、144.91%、294.44%。方差分析表明,T2—T4处理间沟岸土壤有机质含量变化差异显著。上述变化特征表明沟岸土壤有机质含量受葛藤覆被类型(鲜/枯)及覆被物生物量共同影响。枯藤覆被均有助于增加其下沟岸土壤有机质含量,鲜藤覆被则相反。
研究发现,相比裸露沟岸而言,鲜、枯藤覆被沟岸土壤铵、硝态氮含量变化差异明显;鲜藤覆被沟岸土壤铵、硝态氮含量均相对减少;而枯藤覆被沟岸土壤铵态氮含量相对减少(T3处理除外),土壤硝态氮含量则呈现出不同程度的增加。具体表现为:较裸露沟岸,鲜藤覆被沟岸土壤铵、硝态氮含量分别减少了26.63%、12.57%;T2—T4处理土壤铵态氮含量分别减少了25.34%、-25.52%(铵态氮含量增加)、22.08%;T2—T4处理土壤硝态氮含量分别增加了227.75%、161.78%、507.33%。方差分析表明,较裸露沟岸,鲜、枯藤覆被沟岸土壤铵态氮含量变化差异显著;除T1处理外,土壤硝态氮含量变化差异也显著;在枯藤覆被沟岸,T2—T4处理间沟岸土壤铵、硝态氮含量变化差异均显著。上述研究结果表明沟岸土壤铵、硝态氮含量受葛藤覆被类型(鲜/枯)及覆被物生物量共同影响。枯藤覆被有助于增加其下沟岸土壤硝态氮含量,但不利于土壤铵态氮含量的提升;鲜藤覆被下沟岸土壤铵、硝态氮含量均明显减小。
研究发现,相比裸露沟岸而言,鲜、枯藤覆被沟岸土壤有效磷含量均呈现出不同程度的降低,T1—T4处理土壤有效磷含量分别降低了97.63%、78.11%、-47.93%(有效磷含量增加)、21.89%,方差分析表明,较裸露沟岸,鲜、枯藤覆被沟岸土壤有效磷含量变化差异显著,在枯藤覆被沟岸,T2—T4处理间沟岸土壤有效磷含量变化差异显著。表明鲜、枯藤覆被整体上未能提高其下沟岸土壤的有效磷含量,但枯藤覆被可明显增加其下沟岸土壤有效磷含量。
研究发现,相比裸露沟岸而言,鲜、枯藤覆被沟岸土壤速效钾含量均呈现出不同程度的增加。具体地,较裸露沟岸,鲜藤覆被沟岸土壤速效钾含量增幅较小,仅为10.29%;但在枯藤覆被沟岸,沟岸土壤速效钾含量则呈现出倍数级显著增加的规律,且葛藤覆被量越多,效果越明显;T2—T4处理土壤速效钾含量分别提升了213.24%、232.35%、402.94%。方差分析表明,较裸露沟岸,鲜、枯藤覆被沟岸土壤速效钾含量变化差异显著(除鲜藤覆被沟岸T1处理外);在枯藤覆被沟岸,T4较T2、T3处理沟岸土壤速效钾含量变化差异显著。表明鲜、枯藤覆被均有助于提高其下沟岸土壤的速效钾含量,且以提升枯藤覆被沟岸土壤速效钾含量最为明显。
2.3 葛藤覆被对沟岸土壤微生物特性的影响
土壤微生物群落是土壤中最为重要的活体成分,土壤微生物量是指体积小于 5×10-3μm3细菌、真菌等的生物总量,是土壤活性养分的储存库,其数量、组成及其活性是评价土壤质量的重要指标[29]。
覆被状况不同,微生物生长环境如水分、枯落物量和根系分泌物等不同,土壤微生物的数量、组成及活性在某种程度上也存在着较大差别,进而导致土壤微生物群落结构和功能发生改变[30]。表6反映了葛藤不同覆被状况下沟岸表层0—10 cm土壤微生物总PLFA含量及各类微生物PLFA量的相对含量百分比。其中PLFA指的是单位质量的土壤中所包含的所有微生物的量,包括细菌、真菌、放线菌、原生动物等的物质的量,其单位为 nmol/g。
由表6可知,葛藤不同覆被状况下沟岸土壤的总PLFA量之间存在较大差异。鲜、枯藤覆被沟岸土壤的总PLFA量明显高于裸露沟岸(裸露沟岸表层土壤的总PLFA量为3.24 nmol/g,总PLFA量处于最低水平)。其中,鲜藤覆被沟岸(T1)土壤的总PLFA量约为裸露沟岸的10.5倍,总PLFA量最高。枯藤覆被沟岸(T2—T4)土壤的总PLFA 量分别为裸露沟岸的2.6、2.6、6.5倍,土壤的总PLFA量处于裸露沟岸和鲜藤覆被沟岸之间。上述研究结果表明鲜、枯藤覆被均有助于增加其下沟岸土壤的总PLFA量。
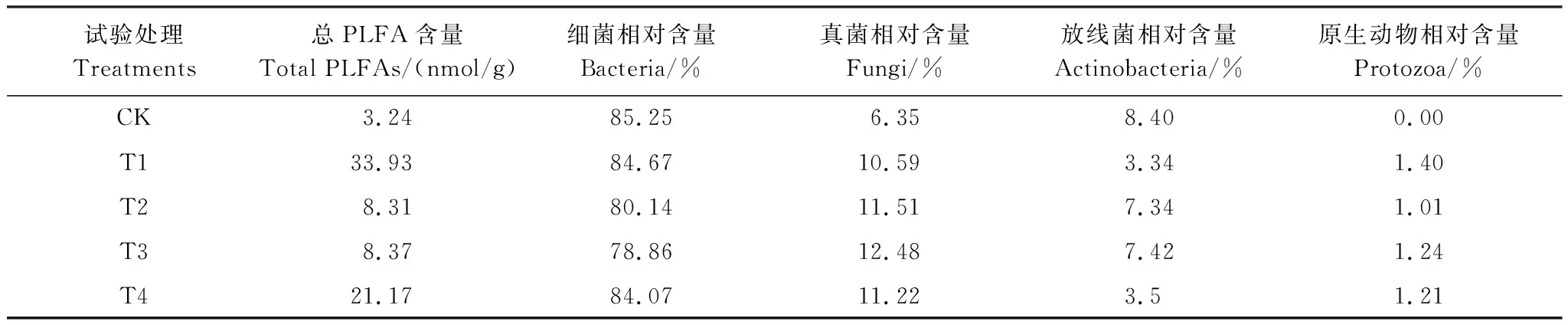
表6 葛藤不同覆被状况下沟岸土壤微生物特性差异
研究还发现,各类土壤微生物(细菌、真菌、放线菌、原生动物)在裸露沟岸和鲜、枯藤覆被沟岸之间存在着不同的变化差异。相比裸露沟岸而言,鲜、枯藤覆被沟岸土壤细菌、放线菌的PLFA相对含量普遍降低,而土壤真菌、原生动物的PLFA相对含量则整体增加。上述变化特征表明沟岸不同种类的土壤微生物对葛藤覆被状况的变化响应不同。
3 讨论与结论
3.1 讨论
本研究葛藤覆被沟岸期间(2006—2016年)不同处理措施下土壤性质均呈现出不同程度的改善,土壤性质的改良效果受葛藤覆被类型(鲜/枯)及覆被物生物量共同影响。与土壤物理性质相比,土壤化学性质及微生物特性更易受葛藤覆被影响而快速被改良。
一般而言,裸露沟岸土壤因无植被生长影响,其容重理应在5种处理中最高,但本研究得到的结果却相对较低。可能的原因在于,尽管裸露沟岸未采取相应的葛藤覆被措施,但该区热辐射强烈,沟岸温度昼夜剧变,表层土壤热胀冷缩,热能疏松表土强烈[9],导致裸露沟岸土壤容重较低。鲜藤覆被沟岸土壤容重为5种处理中的最低值,其降低的原因与葛藤藤茎发达的须根有关,须根扎根土壤后,通过扰动作用疏松了沟岸土壤,降低了土壤容重,出现鲜藤覆被沟岸土壤有机质含量较低,容重也偏低的现象。枯藤覆被沟岸土壤容重略高于裸露沟岸,其可能受以下因素制约,一方面与该地区夏季高温多雨所导致植物残体分解速率快,土壤腐殖化作用弱有关,由于该区土壤腐殖化作用弱,土壤容重的改良也就相对较慢[31];另一方面,在干热暴雨的环境下,表层土壤发生流失,细土颗粒沉积会进入土壤空隙,增加土壤容重[32]。
较裸露沟岸,鲜、枯藤覆被沟岸土壤全盐含量反而增加,这与西北干旱区同类研究结果明显不同[33]。综合分析葛藤不同覆被状况下沟岸土壤全盐含量的变化特征及差异成因:试验沟岸葛藤种植于2006年,沟岸土壤全盐含量为2006—2016年10年间土壤全盐含量变化的阶段性状态量。在裸露沟岸,蒸发最为强烈,盐分表聚现象也最为突出,土壤全盐含量理应最高。但是,裸露沟岸土壤经历着干热河谷雨季逐年周期性的降雨击溅剥蚀和坡面径流侵蚀,陡立沟岸在旱季积累的高盐土壤会在雨季淋失殆尽,因而裸露沟岸土壤全盐含量很低,仅为0.76%。在鲜藤覆被沟岸,葛藤为适应燥热的气候,特别是12:00—16:00强蒸发阶段,葛藤会倾斜叶片以降低蒸发面,期间部分表土裸露,植物蒸腾和土面蒸发同时存在且强烈,土壤水分散失加速,盐分表聚现象明显;在降雨期间,鲜藤叶片又可对降雨产生阻挡,降低了表土剥蚀作用,因而鲜藤覆被沟岸土壤全盐含量较高,达2.28%。在枯藤覆被沟岸,因枯藤存留降低了降雨期间的侵蚀作用,高盐含量土壤得以存留,T2土壤全盐含量(1.26%)较低可能与覆层薄有关,当葛藤覆被物达到一定的厚度后,土壤全盐含量基本恒定,例如试验区T3和T4处理,土壤全盐含量分别为2.27%和2.32%。
较裸露沟岸,鲜藤覆被沟岸土壤有机质、铵态氮、硝态氮、有效磷、速效钾含量大都呈现出不同程度的降低,这可能与藤茎须根扎根土壤以吸收有效养分供给葛藤生长有关。枯藤覆被沟岸土壤上述养分含量整体提升,这与葛藤残体分解后,腐殖质等养分归还土壤有关。需要指出,枯藤亦是由鲜藤发展而来的,在鲜藤覆被沟岸阶段,沟岸土壤养分状况也会出现一定程度的降低,在枯藤覆被沟岸以后,土壤养分状况才得以提升,且土壤养分的主要来源应为沟底葛藤母根根际土壤,具体原因有待下一步研究。
较裸露沟岸,鲜、枯藤覆被沟岸土壤微生物总PLFA量均有所增加,尤其以鲜藤覆被沟岸土壤总PLFA量增加最为明显。鲜藤覆被沟岸土壤总PLFA 量最高可能与葛藤根系分泌物最为密切[30]。枯藤覆被沟岸土壤的总PLFA 量增加的原因则可能与枯藤覆被沟岸后,通过覆层保水、遮蔽降温、提供植物残体等方式提升了土壤含水率、降低了土壤温度、改善了土壤理化性质,尤其是适宜的水热条件加快了葛藤残体分解,提供腐殖质、有效养分等,有益于微生物的生物量和微生物群落活性等的提高。研究还发现,相比裸露沟岸而言,鲜、枯藤覆被沟岸土壤细菌、放线菌的PLFA相对含量普遍降低,而土壤真菌、原生动物的PLFA相对含量则整体增加。即不同种类的土壤微生物对葛藤覆被状况的变化响应不同,这可能与不同土壤微生物对与土壤水热条件、孔隙结构、养分分配比例等的不同有关。
3.2 结论
(1)沟岸土壤性质受葛藤覆被类型(鲜/枯)及覆被物生物量共同影响,与土壤物理性质相比,土壤化学性质(除全盐含量)及微生物特性更易受葛藤覆被影响而快速被改良。
(2)沟岸土壤养分状况受葛藤生长阶段影响。鲜藤覆被沟岸阶段,土壤养分状况呈现出不同程度的降低,枯藤覆被沟岸阶段,土壤养分状况得以明显提升。
(3)鲜、枯藤覆被均有益于增加其下沟岸土壤微生物总 PLFA量,尤其以鲜藤覆被下沟岸土壤总 PLFA量提升最为明显。
(4)整体而言,葛藤覆被可作为沟岸土壤改良的可行方法,它对于提升土壤水分含量、有效孔隙、养分状况、微生物量等均有显著作用。
猜你喜欢
润·文摘(2021年9期)2021-09-22
东坡赤壁诗词(2021年4期)2021-09-05
华南农业大学学报(2021年4期)2021-07-24
煤田地质与勘探(2021年2期)2021-05-23
教学考试(高考英语)(2020年1期)2020-12-25
草堂(2019年10期)2019-11-13
干旱地区农业研究(2019年3期)2019-07-18
红岩(2018年6期)2018-11-16
山东工业技术(2017年21期)2017-11-04
少儿科学周刊·儿童版(2015年1期)2015-07-07
