
氮输入和水淹对短叶茳芏生长、养分及固碳的影响
2018-08-29 11:08张林海曾从盛
生态学报 2018年14期
张林海,曾从盛,仝 川
1 福建师范大学地理科学学院,福州 350007 2 福建省植物生理生态重点实验室,福州 350007 3 福建师范大学湿润亚热带生态地理过程教育部重点实验室,福州 350007
湿地是全球重要的生态系统之一,对全球环境变化异常敏感[1]。同草原和农田等陆地生态系统相比,潮汐盐沼湿地具有较高的固碳和吸存能力,在全球碳循环中扮演重要角色[2]。湿地植物光合固碳作用是湿地碳库的重要来源,然而大部分盐沼湿地生态系统中,氮素常被认为是限制植物光合生长的最重要因素[3]。河口湿地由于地处海洋、大陆和河流的交汇处,是湿地生态系统中特殊的一种类型。近年来,由于人类活动和环境变化导致河口湿地生态系统的氮输入增强,必将对河口湿地生态系统光合固碳过程和碳、氮循环过程产生重要影响[4]。
河口湿地由于特殊地理位置和潮汐作用,湿地植被和土壤常被淡、咸水周期性的淹没[5]。土壤常常为干湿交替状态,造成植物光合生长常受制于干旱或水淹胁迫。水淹状态显著影响植物根系氮的吸收[6]。另外,水分能够对氮的生态效应进行调节,直接影响植物的光合生长。一般认为,氮素可以增强植物的抗旱能力,主要通过降低气孔密度、提高植物的叶绿素含量和叶片束缚水含量,缓解水分匮缺对植物光合的抑制作用[7]。但氮素对改善植物抗水分胁迫能力的作用也会因氮施用量、物种及水分程度而发生改变[8]。河口湿地由于潮汐水淹、径流等的共同作用,外源氮输入后氮的有效性呈现出极大的复杂性,直接影响植物光合固碳能力,进而影响河口湿地生态系统的碳、氮循环过程。目前有关氮输入对河口湿地植物光合生长和固碳的影响已经开展了一些研究,但是多集中在禾本科和藜科类植物[9- 11],针对水淹和氮输入共同作用下河口盐沼湿地莎草科植物的光合生长、固碳的研究较少。
闽江河口湿地处于高氮沉降和高氮营养盐输入地区[12],氮素增加将对植物生长发育、光合和养分循环以及关键生态系统过程产生重大影响[13-14],但是目前关于闽江河口潮汐湿地植物光合生长与固碳对氮输入的响应特征及主要机制的研究还比较缺乏。短叶茳芏为莎草科莎草属多年生草本植物,须根系,地上茎有叶鞘包裹,叶片较少较窄,小穗状花序[15]。在闽江河口湿地,短叶茳芏分布广泛于中、高潮滩,是重要的土著优势种[16]。短叶茳芏虽为草本植物,但是株高可达1.5 m以上,地下根茎发达,根系密集,同时短叶茳芏为C4植物(δ13C丰度实测值为-12.45‰—-13.54‰),具有较强的光合固碳能力。前期的研究表明,闽江河口湿地植物中,短叶茳芏生物量和固碳量较大[17]。同时闽江河口地区的短叶茳芏氮/磷<14,仍主要受氮素养分限制[18]。对于受氮限制、光合固碳能力较强的闽江口河口沼泽湿地的土著植物,氮输入和水淹时长如何改变植物生长、养分和光合固碳能力?开展相关的研究具有重要的意义。本研究以闽江河口土著优势种短叶茳芏为研究对象,通过不同氮输入和水淹条件的模拟实验,对闽江河口湿地短叶茳芏生长特征、碳、氮养分和植物固碳等方面开展研究,旨在进一步探讨氮输入增强等环境变化下河口湿地植物生态适应能力,并为河口湿地的生态环境保护提供参考。
1 研究区域与研究方法
1.1 研究区概况
闽江河口鳝鱼滩湿地(26°01′07″—26°04′34″ N,119°35′51″—119°41′12″ E)是中国东南沿海典型的天然河口潮汐湿地,也是福建省闽江入海口附近面积最大的潮滩湿地。该湿地受潮汐影响明显,水位呈周期性的变化,河口区潮位在冬春季相对较低,夏秋季潮位较高。气候类型为南亚热带海洋性季风湿润气候,温暖湿润,雨热同期,年均温19.6℃,年均雨量1382 mm。鳝鱼滩湿地土壤处于盐质化交替过程,盐分较高,pH呈偏酸性;区内以维管束草本植物占绝对优势,短叶茳芏、芦苇(Phragmitesaustralis)、藨草(Scirpustriqueter)等是该湿地的主要土著优势种,分布广泛[16]。短叶茳芏主要位于该湿地中、高潮滩,高潮滩靠近堤坝处还分布狗牙根(Cynodondactylon)、铺地黍(Panicumrepens)等低矮草本植物。鳝鱼滩湿地由于地理位置独特,动植物多样,是珍稀鸟类的理想栖息地,具有重要的生态保护价值。
1.2 研究方法
1.2.1 实验处理
采样地选择在闽江口五门闸堤外的鳝鱼滩湿地。2014年4月20日,选取短叶茳芏幼苗长势相对一致的地段,小心的采集整株植株及0—20 cm的原位土层,放置到塑料盆中并带回学校实验室。挑选地上植株高度(约高15 cm)和地下根状茎茎节数大体一致的健康植株幼苗,随机移植到直径30 cm、高30 cm的PVC桶中构建中型生态系[19]。植株在每个PVC桶中株数为10株,随机分布;土层深度为20 cm,将边缘土样压实后随机摆放于室外无遮挡、光照充足的地方。用采集的潮水预培养1周,并经过15 d的稳定期后开始进行不同水淹和氮输入处理实验。整个实验持续15个月,至2015年7月15日结束,实验期间植株在自然条件下生长。
根据鳝鱼滩湿地中、高潮滩短叶茳芏植物水淹状况的差异(高潮滩平均水淹时间约为5%—10%,中潮滩平均水淹时间约为40%—50%)。参考徐高峰等[20]的不同水分处理研究方法,本研究设计2种水淹时长处理:T1实验组,在桶侧面15 cm高度处(土深5 cm处)打4个小孔定时放水,每个小孔直径均为1 cm,植物水淹时长约占10%(每天2—3 h);T2实验组,桶侧面底部距地面5 cm处打2个小孔定时放水,植物水淹时间约占50%(每天11—12 h)。根据该区域测定的氮沉降通量9.1 g N m-2a-1[21],参考亚热带农田施氮量15 g N m-2a-1[22],本研究以NaNO3为施氮种类,设置3种氮输入水平:对照CK(0 g N m-2a-1)、N8(8 g N m-2a-1)和N16(16 g N m-2a-1)。将各水平的施氮量平分到12个月,在每月月初和月中,选择晴朗天气各喷施1次。将每次所需的NaNO3溶于1.5 L水中,以溶液的形式喷施于微区土壤表层,结合不同水淹处理进行实验,每种处理3个重复。
1.2.2 实验方法
植株的生长、养分等特征的测定在实验6个月(2014年10月份,植株成熟期)、13个月(2015年5月份,快速生长期)和15个月(2015年7月份,植株生长旺期)后进行。
在各个生长期,每个处理选取6—8株植株测定其株高,同时计数植株的株数。植物密度根据株数和面积推算。采用收获法获取植物地上生物量。植物收获后去除杂物,用蒸馏水冲洗干净,105℃下杀青半小时后在80 ℃条件下烘干至恒重。样品混合后碾磨粉碎,过0.25 mm的筛,测定碳、氮元素的含量。植物碳、氮元素测定采用德国Vario ELIII元素分析仪测定。植物固碳量=植物生物量×碳含量。
1.2.3 数据处理与分析
所有数据使用SPSS 13.0软件进行分析。采用重复测量方差分析(Repeated measures ANOVA)和Duncan多重比较方法比较不同处理间的差异。在进行方差分析前,对数据进行方差齐性检验,本研究中对检验不通过变量(植株株高)的数据进行适当的倒数转换,通过方差齐性检验后再进行方差分析。实验数据均为各指标测定数据的平均值±标准误(SE)。
2 结果与分析
2.1 短叶茳芏生长状况
2.1.1 株高和密度
不同氮输入水平和水淹条件对短叶茳芏不同生长期株高和密度的影响见图1。植物株高和密度的变幅较大,其中株高变化范围为(73.4±1.6)—(114.2±5.5)cm,植物的密度变化范围为(727.7±61.6)—(1464.8±48.8)株/m2。重复测量方差分析表明(表1),不同水淹状况、氮输入水平、生长期对短叶茳芏的株高均有显著影响,T2处理株高极显著高于T1处理,N8、N16处理的植物株高显著高于CK处理,植物成熟季节的株高也显著最高。但是各因素之间的交互作用不显著。不同水淹状况和生长期对短叶茳芏的密度有显著影响,T1处理密度极显著高于T2处理,植物成熟季节的密度也显著最高,但是氮输入对植物密度影响不显著。除氮输入和生长期有交互影响外,其他各因素之间的交互作用并不显著。

图1 不同氮输入和水淹条件处理短叶茳芏株高和密度Fig.1 Dynamic of the height and density in C. malaccensis under different nitrogen addition and submergence treatmentsT1, 水淹处理1, submergence treatment 1; T2, 水淹处理2, submergence treatment 2; CK, 对照, control check; N8, 氮施加量8 g N m-2a-1, nitrogen input 8 g N m-2a-1; N16, 氮施加量16 g N m-2a-1, nitrogen input 16 g N m-2a-1

项目 Items株高Height密度DensitydfMSFPdfMSFP水淹Submergence 10.0013.420.0001397629.9421.630.000氮输入 Nitrogen input20.0015.510.000230956.681.680.199时间 Time20.007.860.0012831609.5345.230.000水淹×氮输入Submergence ×Nitrogen input20.000.070.931221999.541.200.314水淹×时间Submergence×Time20.002.100.137218691.971.020.371氮输入×时间 Nitrogen input×Time40.001.380.262458176.373.160.025水淹×氮输入×时间Submergence×Nitrogen input×Time40.000.990.425442712.302.320.075误差 Error360.003618384.55
df, 自由度, degree freedom; MS, 均方差, mean square;F,F值,Fvalue;P,P值,Pvalue
2.1.2 地上生物量
水淹状况和氮输入对植物地上生物量的影响见表2。重复测量方差分析表明,氮输入水平和生长期对短叶茳芏的地上生物量有显著影响:N8处理的植物地上生物量极显著最高,CK处理极显著最低(F=46.31,P<0.001),植物成熟期的地上生物量也极显著最大(F=93.67,P<0.001)。但是总体上水淹对植物地上生物量影响不显著,表现为实验6个月,T1处理的地上生物量高于T2处理;但是实验13个月和15个月后,T2处理的地上生物量高于T1处理。

表2 不同氮输入和水淹处理短叶茳芏地上生物量/(g/m2)
T1, 水淹处理1, submergence treatment 1; T2, 水淹处理2, submergence treatment 2; CK, 对照, control check; N8, 氮施加量8 g N m-2a-1, nitrogen input 8 g N m-2a-1; N16, 氮施加量16 g N m-2a-1, nitrogen input 16 g N m-2a-1
2.2 短叶茳芏养分含量及固碳量变化
2.2.1 地上碳含量
水淹状况和氮输入对植物地上碳含量的影响见图2。总体上,植物碳含量变幅较小,其变化范围为(384.4±1.1)—(404.2±2.9) mg/g。重复测量方差分析表明(表3),仅有水淹状况差异对植物地上碳含量产生显著影响,其中T2处理地上碳含量显著高于T1处理,氮输入和植物生长期对地上碳含量影响不显著。从各因素交互作用上看,也仅有水淹状况和生长期有显著交互影响,其余因素交互作用不明显。

图2 不同氮输入和水淹条件下短叶茳芏的地上碳含量动态Fig.2 Dynamic of the aboveground carbon content in C. malaccensis under different nitrogen addition and submergence treatments
2.2.2 地上氮含量
水淹状况和氮输入对植物地上氮含量的影响见图3。植物地上氮含量变化范围为(36.5±1.3)—(60.1±2.6) mg/g。重复测量方差分析表明(表3),生长期、水淹状况和氮输入水平对植物地上氮含量都有显著影响:N8处理的植物地上氮含量显著最高,CK处理显著最低;T1处理地上氮含量显著高于T2处理;植物快速生长期地上氮含量极显著最高,成熟期显著最低。氮输入和生长期对地上氮含量有显著交互影响。

图3 不同氮输入和水淹处理短叶茳芏的地上氮含量动态Fig.3 Dynamic of the nitrogen content in aboveground of C. malaccensis under different nitrogen addition and submergence treatments
Table3RepeatedmeasuresANOVAofthecarbonandnitrogencontentinabovegroundofC.malaccensisunderdifferentnitrogenadditionandsubmergencetreatments
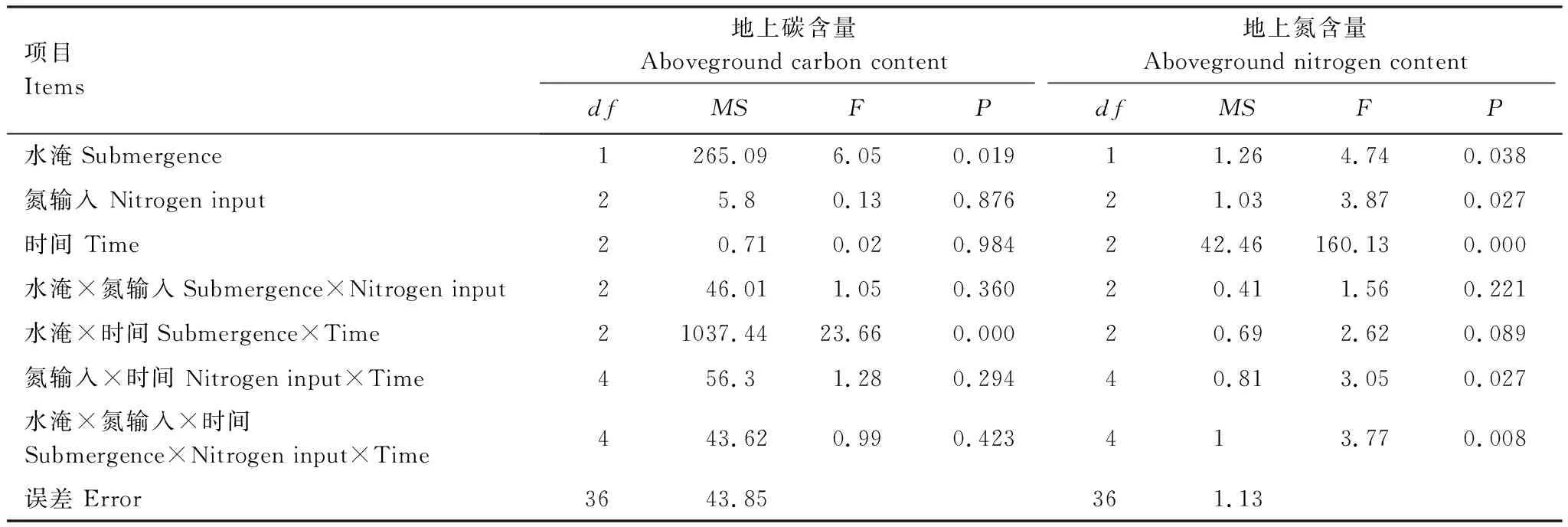
项目 Items地上碳含量Aboveground carbon content地上氮含量Aboveground nitrogen contentdfMSFPdfMSFP水淹Submergence 1265.096.050.01911.264.740.038氮输入 Nitrogen input25.80.130.87621.033.870.027时间 Time20.710.020.984242.46160.130.000水淹×氮输入Submergence×Nitrogen input246.011.050.36020.411.560.221水淹×时间Submergence×Time21037.4423.660.00020.692.620.089氮输入×时间 Nitrogen input×Time456.31.280.29440.813.050.027水淹×氮输入×时间Submergence×Nitrogen input×Time443.620.990.423413.770.008误差 Error3643.85361.13
2.2.3 碳/氮值
2种水淹状态下短叶茳芏地上碳/氮值季节变化比较一致(图4),地上碳/氮值均大于35.0,变化范围为(36.5±1.3)—(60.1±2.6),最低值出现在快速生长期,最高值都出现在生长旺盛期。重复测量方差分析表明,T2处理的植物地上碳/氮值显著高于T1处理(F=8.12,P<0.001),植物在生长旺期碳/氮值极显著最高,快速生长期极显著最低(F=109.84,P<0.001)。氮输入水平及各因素交互作用对地上碳/氮值影响不显著。
2.2.4 地上固碳量
不同水淹和氮输入下短叶茳芏地上固碳量变化情况见表4。重复测量方差分析表明(表5),氮输入水平和生长期对植物地上固碳量影响显著:N8处理地上固碳量极显著高于N16处理,CK处理最低;植物成熟期的地上固碳量也极显著最大。水淹对植物地上固碳量的影响不显著,但是各因素两两交互作用显著。

表4 不同氮输入和水淹处理短叶茳芏地上固碳量/(g C/m2)
3 讨论
水淹状况是湿地植物生长中极为敏感的环境因子[23],湿地植物通过质外体障碍的形成和根系的激素、厌氧多肽等方面的改变以适应水淹环境,还可以在形态和繁殖等方面采取不同的适应对策[24],如通过向氧性生长和增加株高的策略应对水深的胁迫[25]。有研究认为中度水淹时,土壤水分吸力最小,植物吸水耗能最小,吸水容易,可以显著增加植株株高[26],这与本研究的实验结果一致,表明适度增加水淹能导致植株株高的升高。氮是湿地植物生长的必需元素和湿地生态系统的重要限制元素[27],氮输入和水淹对湿地植物生长有较强的耦合作用,对一些耐盐碱植物的研究表明,氮输入+灌水处理均能降低盐胁迫影响,显著提高植物株高和产量[28],本研究中,N8、N16处理的短叶茳芏株高显著高于CK处理(图1),表明氮输入和较长时间水淹对短叶茳芏植株株高也具有叠加促进效应,可能是因为闽江口湿地植物生长仍受氮限制,氮输入加上适宜的水淹可以提高水分利用效率,促进氮素的转化[29]。密度是植物生长的另一个最基本特征,植物密度和株高的变化反映环境发生方向上的逆转或强度上的变化,并体现出种间的竞争和促进关系。本研究中植物密度和株高对氮输入及水淹环境变化的响应并不一致:T2处理株高极显著高于T1处理,而T1处理密度极显著高于T2处理。6个月和13个月,氮输入是影响植株株高的主导因素,植株株高和氮输入也有显著正相关关系,水淹则是影响植物密度的主导因素,短时水淹促进了植物密度增加。15个月后,氮输入是影响植物密度的主导因素,植物密度和氮输入有显著正相关关系,水淹则是影响植物株高的主导因素,较长的水淹促进植株株高增加。植株株高和密度在氮输入与水淹相互作用时的这种互补的响应特征可以在某种程度上指示生境变化和有机体对必需资源的寻求,反映的是植物的投资权衡策略[30]。植物密度和株高的这种响应策略在许多湿地植物的研究都有发现,如周洁和王东[26]对扬子狐尾藻(Myriophyllumoguraense)进行水淹处理后,观测到植株的总生物量、分枝数显著减少,而株高则显著增加;而当湿地植物株高下降时,则倾向于提高种群密度以补偿植株株高降低带来的负效应[31],植物正是通过株高、密度等生长特征不同方式的调节,最终达到耗-益的最大获取,进一步适应环境的变化[25]。
表5不同氮输入和水淹处理植物地上固碳量的重复测量方差分析
Table5RepeatedmeasuresANOVAoftheabovegroundcarbonsequestrationofC.malaccensisunderdifferentnitrogenadditionandsubmergencetreatments
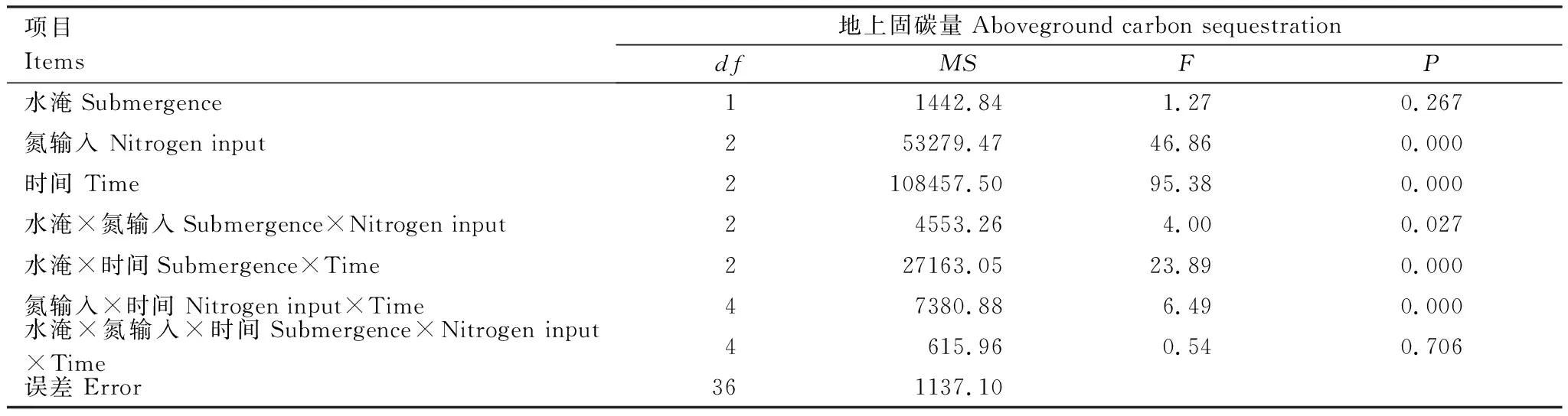
项目 Items地上固碳量Aboveground carbon sequestrationdfMSFP水淹Submergence 11442.841.270.267氮输入 Nitrogen input253279.4746.860.000时间 Time2108457.5095.380.000水淹×氮输入Submergence×Nitrogen input24553.264.000.027水淹×时间Submergence×Time227163.0523.890.000氮输入×时间 Nitrogen input×Time47380.886.490.000水淹×氮输入×时间Submergence×Nitrogen input×Time4615.960.540.706误差 Error361137.10
本研究中,2种水淹处理下,不同氮输入处理植物地上碳/氮值是影响植物固碳的重要因子。实验初期,T1处理植物地上碳/氮值显著较低。地上碳/氮值下降是由于植株体内氮含量增加导致。研究表明,水分不足条件下,植物生长初期叶片较高的氮浓度降低了植物对氮素的需要,此时植物降低生长速率以减小干旱胁迫[32]。也有不一致的结论,如在半干旱环境下,氮输入后植物叶片有较高的碳/氮值,其原因主要有两方面:一是因为干旱导致木质素增加,碳/氮值也相应升高[33]。二是稀释假说认为,碳固定后通过代谢途径稀释了富氮组织对氮的积累[34],因此增加了植物组织的碳/氮值。植株碳/氮值的增加导致更高的氮利用效率,每单位氮能固定更多的碳[35]。但是实验后期不同水淹处理植物地上碳/氮值的差异减小,表明植物在不同生长阶段和各种环境胁迫下不断进行氮素利用与光合碳同化的优化平衡。在水淹或者干旱胁迫条件下,植物对氮素吸收的研究还需要在分子水平和整个物种水平上做进一步的研究[36],同时植物碳/氮值化学计量和光合固碳之间的关系是一个深层次的机理问题,氮输入增强等环境变化对其碳/氮值化学计量的影响还有待进一步深入研究[37]。
水淹对湿地维管束植物光合生长和生物量等有重要影响,湿地植物能通过调节不同植物器官的养分平衡和根系超微结构及根系代谢特性以适应土壤水分和养分的变化[38],取决于物种的特异性[39]和细胞渗透势调节机制[40]。一般认为,较长的潮水淹没导致植物缺氧与光合降低,进而抑制植物生长[41],如当滞水和淹水时间达到3周以上时,柳叶菜(Epilobiumhirsutum)的生物量减少[42]。对滨海盐沼互花米草(Spartinaalterniflora)施氮后表明,地上部生物量在施氮条件下显著增加,且在土壤处于间歇淹水状态时表现更为明显,而持续淹水状态对互花米草生长和光合具有抑制作用[11]。本研究中,实验6个月后,T1处理的地上生物量显著高于T2处理,这是由于T2处理中较高的水淹深度和水淹时间导致短叶茳芏光饱和点(LSP)、叶绿素相对含量(SPAD值)显著降低,蒸腾速率(Tr)升高造成[43],因此植物的生物量和氮含量也更低。氮输入也显著影响植物生长和固碳的积累。多数研究认为氮输入增加了植物生物量和固碳量,如研究表明氮输入促进沼泽湿地茭草(Zizaniacaduciflora)地上生物量积累[44],也明显促进了小叶章(Calamagrostisangustifolia)的株高和叶面积,显著增加了碳的生物量积累[10]。本研究中,氮输入也显著提高了短叶茳芏的地上生物量和固碳量。这些研究表明在异质环境中,湿地水生植物能通过自身调节机制及生长策略的调整产生适应性,拓宽其生态幅,这对湿地植被恢复和生物多样性维持有重要意义[45]。本研究在氮输入条件下,较短时长的水淹处理植物的密度、氮含量和前期固碳量较高,说明短叶茳芏在氮输入下,可以通过生长、养分和光合固碳量的调整,补偿由于干旱和水分降低等生长不利因素的影响。考虑到短叶茳芏较广的适盐性和水淹环境适应能力,在持续氮输入条件下,闽江河口湿地短叶茳芏有向高潮滩拓展的可能,这有利于闽江河口湿地土著优势种的维持和恢复。
猜你喜欢
汽车实用技术(2022年16期)2022-09-03
今日农业(2020年19期)2020-12-14
小哥白尼(军事科学)(2018年3期)2018-06-15
上海农业学报(2017年4期)2017-04-10
上海农业学报(2016年5期)2016-02-10
上海农业学报(2016年5期)2016-02-10
中国科技信息(2015年2期)2015-11-16
河南科技(2015年3期)2015-02-27
中学生理科应试(2014年2期)2014-04-23
植物营养与肥料学报(2014年1期)2014-03-11
