
2个红麻品种在改良铅锌矿渣下的耐性研究
2018-08-29 01:46龚紫薇陈永华陈基权张倩妮欧丹玲柳俊
中国麻业科学 2018年4期
龚紫薇,陈永华*,陈基权,张倩妮,欧丹玲,柳俊
(1.中南林业科技大学环境科学与工程学院,长沙410004;2.中国农业科学院麻类研究所,长沙410205)
铅锌尾矿是一类Pb、Zn含量极高且对植物的生长产生胁迫的生境,采矿后破坏了矿区大部分的植被与耕地,并产生大量的矿业废物,对土壤质地和结构产生严重的影响。矿业废物中重金属含量较高,是主要污染源之一[1-2],其导致的重金属污染已引起国内外广泛关注[3-6]。而选用重金属耐受力强,生物量大,速生的植物对修复重金属污染严重的铅锌尾矿库起到了关键性的作用。
植物对重金属的抗性通常表现为对重金属的避性和耐性,耐性又包括金属排斥和金属富集两个方面[7]。选择性吸收、根部滞留、区域化作用等都是植物耐受重金属毒害的策略[8],亚细胞分布可为研究植物对重金属的解毒和生物富集机制提供重要信息[9]。已有研究[10-11]表明,植物吸收重金属后,会将其以不同的化学形态存在于体内,从而限制重金属在体内的迁移,达到缓解毒性的目的,因此,重金属在植物体内的存在形态对揭示植物对重金属的耐性也有重要意义。
红麻(Hibiscus cɑnnɑbinus)由于具有生长迅速、较强的耐干旱、耐涝性以及巨大的生物产量等特性[12],对多种重金属也具有较强的耐性[13-14],且红麻作为非食用纤维经济作物,可避免重金属通过食物链对人体造成伤害,又可以获得较理想的经济效益,有望成为植物修复尾矿库重金属污染的一类有价值的经济作物。
本课题组前期研究发现,红麻品种闽红362和T14相对于同批筛选品种,能在改良铅锌尾矿渣中良好生长,对高浓度复合重金属污染表现出较强的耐性。在此基础上,本研究拟进一步分析这两个红麻品种对Pb、Zn的富集能力及Pb、Zn在植物体内的亚细胞分布和化学形态特征,以揭示其对Pb、Zn复合污染的耐性及其耐性机制。
1 材料与方法
1.1 试验材料
闽红362和T14种子由中国农业科学院麻类研究所提供,铅锌矿渣来自湖南省郴州市某铅锌尾矿库,其基质本底值见表1,泥炭土购买于长沙红星花卉市场(pH值约为6.45,有机质含量约为90 g/kg)。
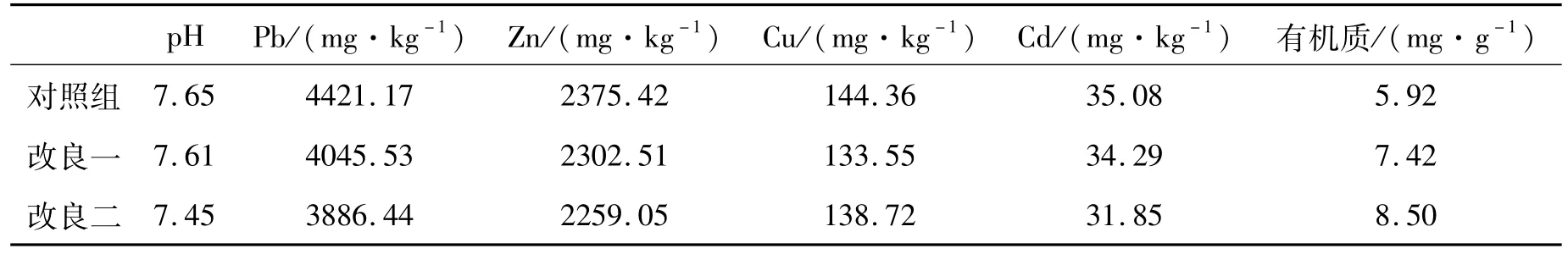
表1 基质本底值Tab.1 Matrix background value
1.2 试验设计
盆栽花盆口径为30 cm,储量为8 kg,泥炭土改良剂设置3个梯度,分别为对照组(CK):100%铅锌矿渣+少量磷肥;改良一(T1):15%泥炭土+85%铅锌矿渣+少量磷肥;改良二(T2):30%泥炭土+70%铅锌矿渣+少量磷肥,3个平行,4月将红麻种子先播种于非重金属污染土壤中,待幼苗长至10~15 cm时将其移栽到已配置好的矿渣改良基质中,10月成熟后收获。
1.3 样品采集
将收获植株先用自来水冲洗,再用去离子水冲洗干净,吸干表面水分。每个处理的植物样品分为根、茎和叶这3个部位,分别用于测定重金属全量、亚细胞组分和化学形态,剩余样品保存于-4℃冰箱内备用。
1.4 指标测定
(1)植物对Pb、Zn的富集与转移能力指标测定
植物样品重金属含量采用火焰原子吸收分光光度计测量,采用“硝酸-高氯酸体系湿法”消解。重金属富集系数和转运系数计算公式如下:
富集系数(BCF)=植物体内重金属含量/根系土壤中重金属含量
转运系数(TF)=植物地上部分重金属含量/根部重金属含量
(2)植物亚细胞组分的分离和化学形态分析
采用差速离心法分离不同的细胞组分[15]。
(3)植物体内Pb、Zn的化学形态分析
采用化学试剂逐步提取法提取植物体内的重金属化学形态[16]。
1.5 数据处理
采用SPSS 22软件分析数据。
2 结果与分析
2.1 红麻对Pb、Zn的富集与转运分析
由红麻植株各部分Pb、Zn含量(表2)可知,Pb、Zn主要储存于根部,但不同改良剂浓度处理,红麻品种各部位的Pb、Zn含量存在差异。随着改良剂浓度的增加,两个品种红麻体内Pb、Zn含量均呈现对照组>改良一>改良二的规律,但地上部Zn含量与之相反,可能原因是Zn元素是植物生长的必需元素,参与植物体内重要酶的合成,促进植物体内氮素的代谢,因此含量更高。
从植物的富集系数(表2)可以看出,两红麻品种对Pb、Zn的富集系数存在差异,对Zn的富集能力大于对Pb的富集能力。随着改良剂浓度的增加,植物Pb、Zn的富集系数总体上呈现对照组>改良一>改良二的规律。各处理下闽红362对Pb的富集能力均大于T14,而Zn的富集系数与之相反。
从植物的转运系数(表2)可以看出,两红麻品种对Zn的转运系数大于对Pb的转运系数,且随着改良剂浓度增加而增加,植物Pb的转运系数在各处理间变化不明显,Zn的转运系数呈现对照组<改良一<改良二的趋势。
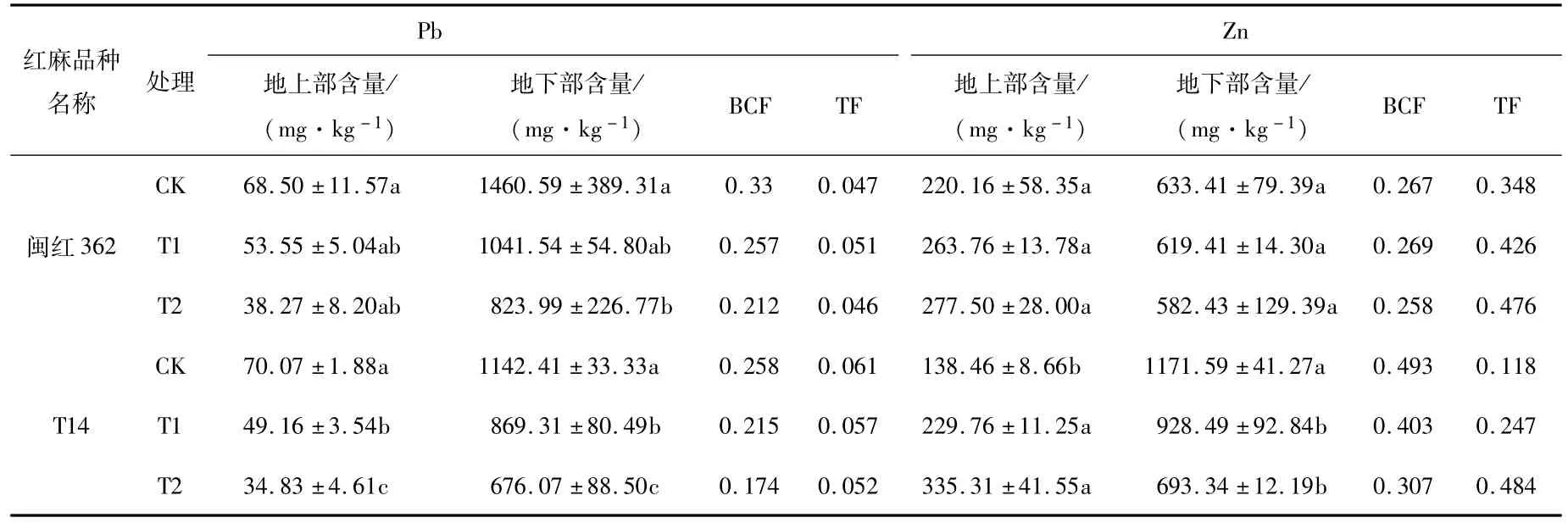
表2 植物各部分Pb、Zn含量及其富集、转运系数Tab.2 Pb and Zn contents in various parts of plants and their enrichment and transport coefficients
2.2 红麻体内Pb、Zn的亚细胞分布特征
由植物体内Pb的亚细胞分布(图1)可知,两种红麻植物叶片中的Pb主要分布于细胞器和细胞壁,占总量的75%~86%,添加改良剂后,细胞器中Pb所占比例下降;茎中的Pb主要分布在细胞壁中,占总量的58%~72%,加入改良剂后,细胞器中Pb所占比例有不同程度的下降;根中的Pb主要集中在细胞壁内,占总量的71%~82%,添加改良剂后,闽红362细胞器和细胞液中Pb所占比例下降,而T14细胞液中Pb所占比例上升。
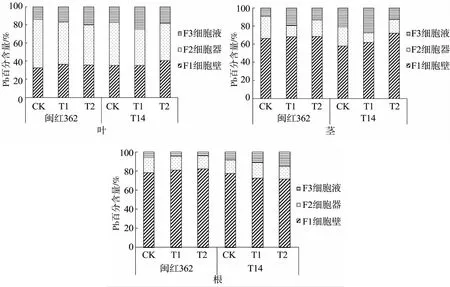
图1 植物各部位Pb的亚细胞分布Fig.1 Subcellular distribution of Pb in various parts of plants
由植物各部位Zn的亚细胞分布(图2)可知,两种红麻植物叶片中的Zn主要分布在细胞壁和细胞液中,占总比例的92%~97%,改良剂的加入降低了闽红362细胞器内Zn的占比,但提高了T14细胞器内Zn的占比。
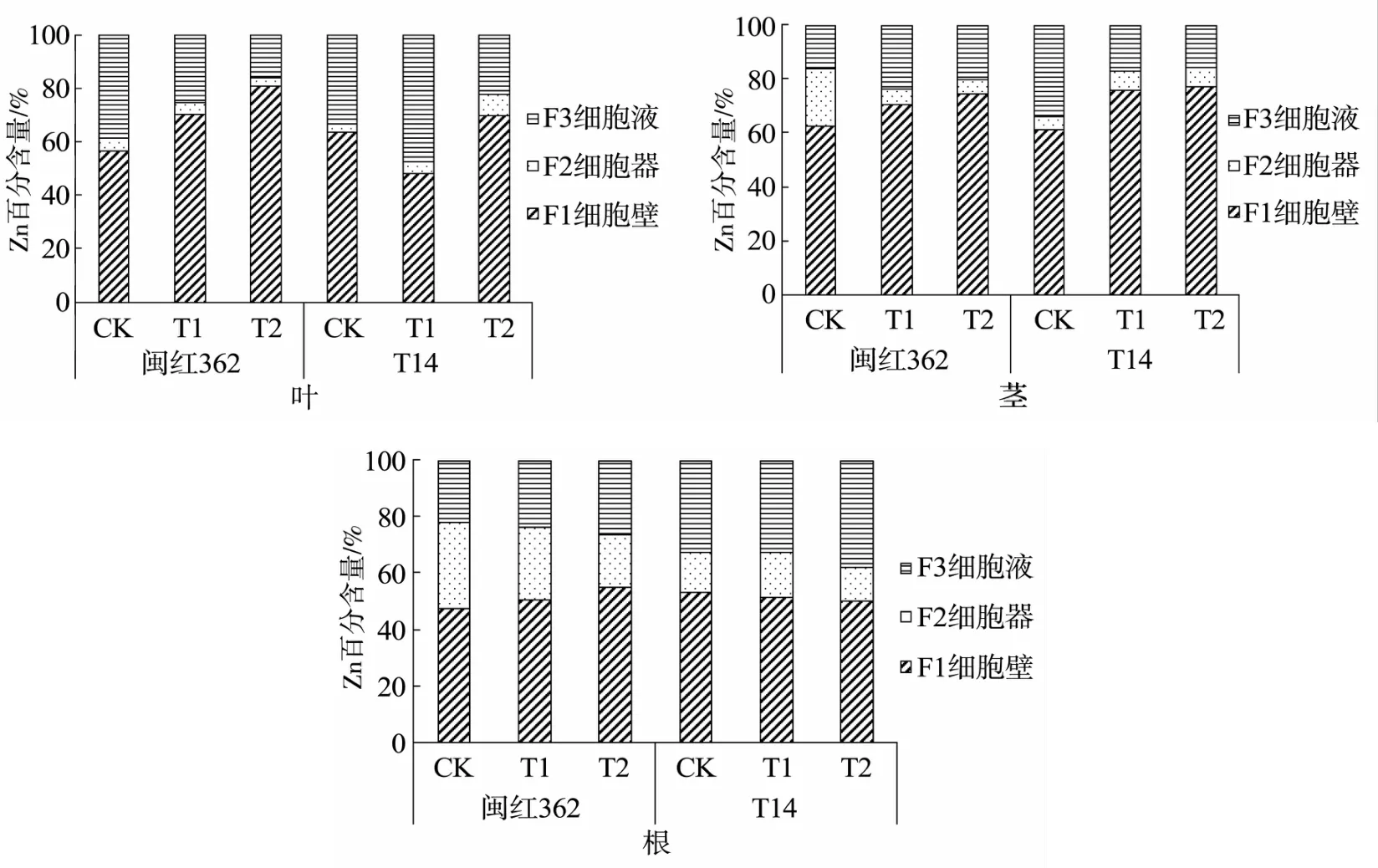
图2 植物各部位Zn的亚细胞分布Fig.2 Subcellular distribution of Zn in various parts of plants
除闽红362对照组,两种红麻植物茎中的Zn主要分布在细胞壁和细胞液内,占总比的93%~95%,加入改良剂后,其细胞壁的占比均有上升,说明改良剂的加入增强了细胞壁对Zn的滞留作用。但其对细胞器和细胞液的影响不同,闽红362细胞器占比下降,细胞液占比上升,T14与之相反。闽红362根中的Zn主要分布在细胞壁中,细胞壁和细胞液占比呈现对照组<改良一<改良二的规律,T14主要分布在细胞壁和细胞液内,各部分占比在加入改良剂后变化不大。
2.3 红麻体内Pb、Zn的化学形态特征
从植物体内Pb化学形态分布(图3)来看,两红麻品种叶片中Pb主要以去离子水提取态和氯化钠提取态存在,其次是乙醇提取态;茎中的Pb主要以醋酸提取态存在,占总比例的66%~82%,活性较高的乙醇提取态和去离子水提取态之和仅占不足10%,且随改良剂加入量的增加而降低,醋酸提取态则有所升高;在对照下,两种植物根部的Pb主要以氯化钠提取态和去离子水提取态存在,随着改良剂的加入,活性较高的乙醇提取态和去离子水提取态所占比例有不同程度的下降,使得氯化钠提取态成为主导形态,闽红362醋酸提取态和盐酸提取态占比有所下降,而T14则相反。
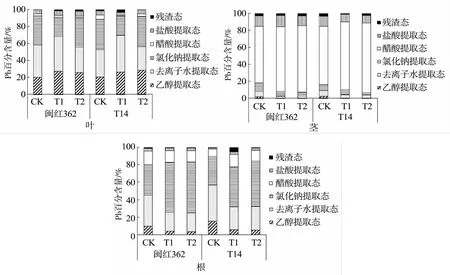
图3 植物体内Pb化学形态分布Fig.3 Distribution of chemical speciation of Pb in plants
从植物体内Zn化学形态分布(图4)来看,两红麻品种茎、叶内的Zn主要以氯化钠提取态和去离子水提取态存在。改良剂加入后,在叶片中,活性较强的乙醇提取态和去离子水提取态表现出对照组<改良组的规律,活性较弱的氯化钠提取态和醋酸提取态则表现为对照组>改良组;而在茎部,乙醇提取态和去离子水提取态则表现出对照组>改良组,氯化钠提取态表现为对照组<改良组;与地上部不同,两红麻品种根部Zn主要以醋酸提取态存在,占比达45%~76%,改良剂的加入增加了活性较强的乙醇提取态和去离子水提取态的占比。
3 讨论
3.1 两红麻品种对Pb、Zn的富集与转运能力
从重金属元素本身特性看,Zn是植物生长必需的营养元素,对植物的生长发育起着重要作用,而Pb是植物生长非必需元素,不管是否为必需元素,只要植物体内重金属含量超过一定的临界值就会对植物生长产生胁迫,造成植物体内代谢活动紊乱,使植物生长发育受到抑制,甚至死亡[17]。本研究基质中Pb、Zn含量分别高达3886.44~4421.17 mg/kg和2259.05~2375.42 mg/kg,远超过植物的正常耐受阈,供试的两品种红麻均成活下来,说明其具有一定的耐性,且表现出一定的Pb、Zn富集能力,这与张富运[18]对红麻的研究结果类似。
不同品种植物以及不同植物器官对重金属的吸收、富集能力不同[19-21]。本试验发现,Pb、Zn主要分布在两红麻品种的根部,说明红麻对重金属的耐性机制主要是排除机制[22],可能是因为根直接与土壤接触、吸收,从而在根部产生高浓度的重金属含量[23]。另外,土壤中的 Pb不易被植物吸收,其可溶性、生物可利用性、移动性较差[24],重金属元素Zn虽然是植物所必需的微量元素之一,但该试验生长基质中Zn含量远超过植物所需,植物吸收 Pb、Zn后将其保留在根部,限制有害或多余的Pb、Zn离子由根部向植物茎叶转移,减少其对植物体的毒害[25]。
两红麻品种对Pb、Zn的富集与转运系数均未达到超富集植物的标准,但其对Pb、Zn的富集与转运能力表现出差异。可能是因为Zn是植物生长所必需的元素,植物对Zn的转运能力远超过对Pb的转运能力,因而植物地上部Zn的含量及其转运系数随改良剂浓度的增加而增加。不同品种对同种重金属元素的富集能力存在差异,本试验中闽红362对Pb的富集能力强于T14,而T14对Zn的富集能力强于闽红362,周航[26]对玉米的研究也出现过类似的结果。
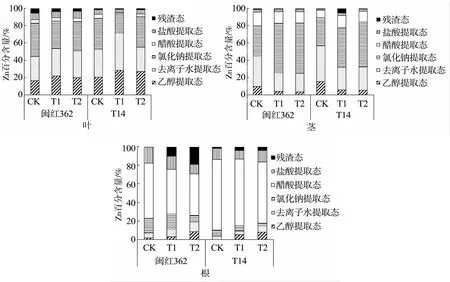
图4 植物体内Zn化学形态分布Fig.4 Distribution of chemical speciation of Zn in plants
3.2 Pb、Zn在红麻体内的亚细胞分布
重金属在细胞壁的沉淀以及细胞内的区隔化分布是植物内部解毒的两个重要途径[27-28]。朱光旭等[29]的研究表明,生长在铅锌尾矿污染农田上的3种菊科植物体内的Pb和Zn主要分布在液泡可溶组分和细胞壁中,而在细胞器中的分布较少,与本研究的结果类似。重金属离子带正电,因而很容易在带负电的细胞壁处被大量络合而沉积[30]。另外,植物细胞壁中含有蛋白质和多糖以及大量亲金属离子的配位基团,容易固定重金属离子[31-32]。当细胞壁结合的金属离子达到饱和点时,进入细胞内的金属离子被转运到液泡中储藏起来,液泡中的多种蛋白质、有机酸、有机碱等物质,都能与重金属结合,从而降低金属离子的毒性[33]。Cobbett[34]研究认为,重金属在非代谢活性组织中的分布可减轻或避免其对功能性结构单元的损伤和代谢过程的干扰。两红麻品种体内的Pb都主要分布在细胞壁组分中,细胞壁对重金属的固持作用是其耐受Pb的重要机制。而其体内的Zn则主要存在于细胞壁和细胞液,可见,细胞壁和液泡对Zn的区域化作用是红麻的耐Zn机制之一。
随着基质中Pb、Zn浓度的增加,闽红362根部细胞壁的分配比例下降,而T14有所增加,这可能是由于T14细胞壁中多糖、蛋白质等对Pb、Zn的沉淀、络合数量及能力要比闽红362强[35]。
3.3 Pb、Zn在红麻体内的化学形态
重金属在植物中的化学形态影响着金属离子在植物体内的迁移、活性以及对植物的毒性[36]。植株体内吸收的重金属有着不同的化学形态,随着乙醇、去离子水、氯化钠、醋酸和盐酸等提取剂的极性增强,所提取出的重金属在活性、毒性和迁移能力上不断降低[37]。茶树银霜品种根部的Pb主要以氯化钠提取态为主[38],小麦茎叶部中Pb以氯化钠提取态、醋酸提取态和去离子水提取态存在[20],皖景天在高浓度Zn胁迫下,根部的Zn主要以醋酸提取态存在[39]。与之类似,本试验中两红麻品种体内Pb、Zn主要以氯化钠提取态和醋酸提取态存在,说明Pb、Zn在红麻体内主要以蛋白质结合或吸附态及果胶酸盐的形式存在,可能与细胞壁和细胞膜中果胶酸和蛋白质等的固定作用有关;其次是以磷酸盐和草酸盐的形式存在,而这部分移动性较差形态的重金属可能与液泡中有机酸的络合作用有关。
随着改良剂浓度的增加,植物根部Zn的乙醇提取态和去离子水提取态的占比上升,而乙醇和去离子水提取态的迁移能力最强,这也解释了为何植物地上部Zn的转运系数随改良剂添加量的增加而增加。
4 结论
(1)红麻能在高浓度Pb、Zn复合污染的矿渣基质中存活,但Pb、Zn在不同品种及不同部位的分布存在差异,植物对Zn的富集与转运能力高于对Pb的富集与转运能力,且随着改良剂浓度的增加,Zn的转运系数呈增大的趋势。因此,泥炭土对铅锌矿渣的改良是成功的,但在实地修复中,建议至少应对修复植物根际矿渣进行改良处理,还可尝试对铅锌矿渣与泥炭土的比例作小范围的调整。
(2)从红麻植物不同部位Pb的亚细胞分布来看,Pb主要存在于细胞壁组分中,改良剂的加入降低了Pb在细胞器的占比;从不同部位Zn的亚细胞分布来看,Zn主要分布于植物的细胞壁和细胞液,改良剂的加入使闽红362茎部Zn在细胞液的占比上升,而在T14中则呈现下降趋势。
(3)Pb和Zn在植物体内主要以氯化钠提取态和醋酸提取态存在,改良剂的加入,使在植物体内活性较强的Pb化学形态占比下降,而根部Zn活性较强的化学形态占比上升。
猜你喜欢
江苏农业科学(2022年19期)2022-10-28
中国农业科学(2022年16期)2022-09-19
山西农业科学(2022年8期)2022-08-12
氯碱工业(2021年6期)2021-12-25
中国土壤与肥料(2021年5期)2021-12-02
河南建材(2021年2期)2021-03-11
三农资讯半月报(2020年18期)2020-10-14
石油化工应用(2020年7期)2020-08-08
数码世界(2018年1期)2018-12-23
浙江工业大学学报(2017年5期)2018-01-22
