
室内三种富营养模拟条件下水流调控对羊栖菜和石莼生长及生物质合成的影响
2018-08-25 06:35:58WERNBERGThomasDEBETTIGNIESThibaut潘耀茹周朝生黄志行吴嘉平
浙江农业学报 2018年8期
于 琰,WERNBERG Thomas,DE BETTIGNIES Thibaut,3,潘耀茹,李 超,林 芳,周朝生,黄志行,吴嘉平,肖 溪,*
(1.浙江大学 海洋学院,浙江 舟山 316021; 2.西澳大学 植物生物学学院,澳大利亚 珀斯 6009; 3.国家自然历史博物馆,法国 巴黎 75005; 4.浙江大学 环境与资源学院,浙江 杭州 310058; 5.浙江海洋水产养殖研究所,浙江 温州 325005)
由人类活动引起的近岸海域富营养化已成为全球海洋环境研究的焦点之一[1-2]。而大型海藻养殖被认为是一种环境友好型的近海富营养化生物修复方法[3-5]。大型海藻作为海洋生态系统重要的初级生产者,可为海洋生物提供氧气、食物和栖息地[6],缓冲海浪对海岸的冲击侵蚀,同时移除过量营养盐[7-10]。研究表明,1 hm2养殖的大型海藻可移除相当于17.8 hm2和126.7 hm2中国近岸海域海水中的氮和磷[11]。此外,养殖的大型海藻也是许多亚洲国家和地区重要的食物来源[12-13]。目前我国大型海藻养殖产量占全球总产量的47.9%,占全球总产值的47.4%[14]。因此,相较于物理修复、化学修复等其他修复技术,大型海藻养殖修复富营养化水域不失为一种最经济安全有效的原位生物修复技术[8, 15-17]。然而,如何提高营养盐生物修复效果和以细胞色素和富氮化合物为代表的生物质含量是该生物修复技术所面临的难点。营养盐供给是影响海藻生长的重要因素[18],之前的大部分研究都在低浓度或中浓度营养盐条件下开展[19-20],但在富营养化水域中营养盐则不再成为限制因素。除该因素外,水流交换也在藻体生长时发挥重要作用[21]。水流速度增加能够减小藻体表面扩散边界层厚度从而加速养分和CO2的供应与吸收,促进其生长[21-23],同时加速藻体代谢产物如O2、OH-、过量的过氧化氢和卤化有机物等物质的移动[24]。当水流交换率由2 vol·d-1提高到8 vol·d-1时,硬石莼(Ulvarigida)生长率提高33.3 %[25]。当水流交换率由5 vol·d-1提高至10.2 vol·d-1时角叉菜(Chondruscanaliculatus)产量明显提高[26]。巨型海带(Macrocystispyrifera)的无机氮吸收速率在水流速度达到2~6 cm·s-1时达到最大[21, 27-28],裙带菜(Undariapinnatifida)则可能会更高(>10 cm·s-1)[29]。而低水流速度则可能通过降低传质限制从而减弱藻体净光合作用效率,研究表明,当水流速度小于1 cm·s-1时,龙须菜(Gracilariaconferta)光合作用受到限制[24]。
本研究选取两种我国近岸海域较为常见的大型海藻——羊栖菜(Sargassumfusiforme)和石莼(Ulvapertusa)作为实验藻种,在实验室环境中调控水流交换率,探究3种营养化程度海水应用情境下两种大型海藻的生长规律;并检测藻体细胞色素(叶绿素a、胡萝卜素)和富氮化合物(可溶性蛋白、可溶性碳水化合物)浓度的变化情况。并利用自行设计的旋转养殖系统,进一步探究调控水流速度对两藻种生长的影响。旨在明确富营养化条件下水流改变对两种大型海藻生长和生物质积累的作用,为今后富营养化海域大型海藻生物修复选种工作提供实验依据,以期优化基于大型海藻的富营养化海域和海水养殖污水净化工艺,提高其营养盐移除效果。
1 材料与方法
1.1 实验材料
2015年9月至11月,在我国浙江省温州市洞头县,浙江水产养殖研究所洞头基地(27°51′42″, 121°11′06″)养殖海域采集褐藻羊栖菜(Sargassumfusiforme)和绿藻石莼(Ulvapertusa)。用过滤后的自然海水清洗藻体表面,去除泥沙和附生动植物。暂养在盐度28‰的过滤自然海水中,用冷水机将温度控制为18 ℃(HS-90A,广东海利有限公司,中国),荧光灯提供光照,强度为120 μmol photons·m-2·s-1,光照周期为12 h/12 h(L/D)。
1.2 实验设计
根据目前我国近岸富营养化状况[30]并设想极端情况,在自主设计实验体系下,模拟近岸富营养化海水、养虾池废水及极度富营养化海水3种环境,对3个110 cm×100 cm×40 cm大玻璃水缸中过滤海水的营养盐进行加富,使最终铵盐和磷酸盐浓度分别达到25 μmol·L-1与2.5 μmol·L-1,50 μmol·L-1与5.0 μmol·L-1,100 μmol·L-1与10.0 μmol·L-1,在每个大缸分别放入5个20 cm×36 cm×25 cm小玻璃水缸并在每个小水缸中投放暂养两天且健康程度与生长状况基本一致的藻体,用小功率水泵(HJ-2200,森森有限公司,中国)与液体转子流量计(LZB-10,余姚银环流量仪表有限公司,中国)调控使其达到设定的水流交换率0、25、50、100及200 vol·d-1。光照强度、光照周期与盐度等其他条件与暂养条件保持一致,每2 d更换一次营养盐加富海水。每个处理重复6个平行样,培养周期为14 d。
中试同样以暂养2 d且健康程度与生长状况基本一致的羊栖菜、石莼藻体作为实验材料,在极度富营养化海水情境的大玻璃水缸中将藻体固定于自主设计的旋转养殖系统最外圈处,半径1/2处和半径1/3处,通过转速调整按钮将转盘转动线速度达到0.3 m·s-1,则对应半径1/2处和半径1/3处的转动线速度则为0.15和0.10 m·s-1。光照强度、光照周期与盐度等其他条件与暂养条件保持一致,每两天更换一次营养盐加富海水。每个处理重复6个平行样,培养周期为12 d。
1.3 生理生化指标测试
1.3.1 比生长速率
在藻类生理生化研究领域中,比生长速率(specific growth rate,SGR)是衡量大型藻类生长状况的经典指标之一。称量前用吸水纸或者毛巾轻轻擦拭藻体表面,使藻体表面干燥程度尽量保持一致,称量藻体实验前后的鲜质量来计算比生长速率[31]。
1.3.2 光合色素
光合色素(photosynthetic pigment)指标包括叶绿素a (chlorophyll a, Chl a)和胡萝卜素(carotene, Car),其含量的测定均采用丙醇提取法[32]。采用分光光度计(T6,上海元析有限公司,中国)在450 nm、666 nm和730 nm波长处测定吸光值。
1.3.3 可溶性蛋白
可溶性蛋白含量(Soluble Protein, SP)的测定采用考马斯亮蓝G-250法[33]。用分光光度计在595 nm波长测定吸光值。
1.3.4 可溶性碳水化合物
可溶性碳水化合物(soluble carbohydrate,SC)含量测定则采用苯酚—硫酸法[34]。用分光光度计在490 nm波长处测定吸光值。
1.4 统计分析
利用Microsoft Excel 2007、SPSS 20.0、R语言对每组的6个平行样进行分析,所有处理组进行双因素方差分析,设置显著水平为P<0.05,并采用Tukey’s HSD方法检验各处理组显著性差异。
2 结果与分析
2.1 比生长率
图1中在养虾场废水应用情境下羊栖菜比生长率明显高于其他水流处理组,且当水流交换率为200 vol·d-1达到最大值(4.34 ± 0.11)%·d-1,而在极度富营养化海水应用情境下出现最小值(0.67 ± 0.29)%·d-1。表1双因素方差分析结果显示,水流交换率和富营养化应用情境两因素的交互作用对羊栖菜比生长率影响显著(P<0.05),富营养化应用情境即营养盐浓度对羊栖菜比生长率有极显著影响(P<0.01)。所有培养条件下,石莼的生长均明显优于羊栖菜。养虾场废水、极度富营养化海水各处理组石莼生长状况均优于近岸富营养化海水应用情境下。当水流交换率为100 vol·d-1,中、高浓度营养盐即养虾场废水、极度富营养化海水条件下石莼比生长率分别为(6.31±1.42)%·d-1、(8.00±0.79)%·d-1,明显高于其他处理组。富营养化应用情境和水流交换率分别对石莼比生长率有极显著影响(P<0.01)和显著影响(P<0.05)。
2.2 生化指标
如图2所示,羊栖菜的所有处理组中,水流交换率为0 vol·d-1、养虾场废水模拟环境下叶绿素a浓度达到最高(0.327±0.017)mg·g-1,其他处理组其含量维持在0.158~0.309 mg·g-1。类胡萝卜素浓度则在水流交换率为200 vol·d-1、养虾场废水模拟环境下达到最高(0.054±0.018)mg·g-1,其他组分其含量维持在0.020~0.037 mg·g-1。表2双因素方差分析结果表明,水流交换率、富营养化应用情境及两者交互作用对羊栖菜类叶绿素a和胡萝卜素浓度均无显著性影响(P>0.05)。极度富营养化海水环境下可溶性蛋白浓度最高,养虾场废水环境下次之,近岸富营养化海水条件下最低。富营养化应用情境对羊栖菜可溶性蛋白浓度有显著性影响(P<0.05)。可溶性碳水化合物浓度在养虾场废水环境、水流交换率为50 vol·d-1时达到最大(26.460 ± 7.302) mg·g-1,在极端富营养化海水情境、水流交换率为100 vol·d-1时最小(5.549 ± 0.373) mg·g-1,与其他处理组的差异显著(P<0.05)。如图3所示,中浓度营养盐即养虾场废水环境条件下的Car/Chla比值明显高于其他两种应用情景,且随水流交换率的增加而增大。就SC/SP而言,近岸富营养化海水、养虾场废水环境下其比值明显高于极端富营养化海水环境,富营养化应用情境仅对SC/SP比值有显著性影响(P<0.05),而水流交换率及两者交互作用对两比值均无显著性影响(P>0.05)。
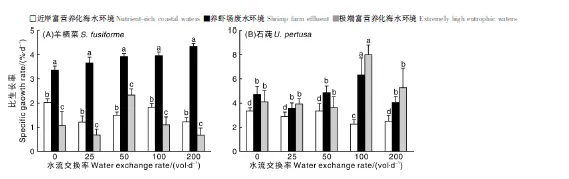
不同处理间没有相同小写字母表示差异显著(P<0.05)。The bars with different lowercase letters showed the significant difference (P<0.05).

表1 应用情境,水流交换率及两者的交互作用对羊栖菜和石莼比生长率影响的双因素方差分析Table 1 ANOVA analysis of the effect of application scenarios, water exchange rate, and their interaction on specific growth rate (SGR) of Sargassum fusiforme and Ulva pertusa
图4中石莼的叶绿素浓度和类胡萝卜素浓度呈现出相同的趋势,养虾场废水、极端富营养化海水环境下,两物质浓度均高于近岸富营养化海水环境下的两者浓度,养虾场废水条件下两物质浓度均值分别为(1.089±0.083)mg·g-1、(0.319±0.007)mg·g-1,极端富营养化海水条件下两物质浓度均值分别为(1.082±0.015)mg·g-1、(0.454±0.004)mg·g-1。可溶性蛋白则呈现与前两者相反规律,近岸富营养化海水、养虾场废水环境下其浓度高于极端富营养化海水环境,在养虾场废水环境、水流交换率为25 vol·d-1条件下最高达(7.367±0.160)mg·g-1,养虾场废水环境、水流交换率为0 vol·d-1条件下最低为(5.247±0.005)mg·g-1。而可溶性碳水化合物浓度在处理组中无明显规律。表3显示营养盐浓度即海水富营养化应用环境对4种物质浓度均有显著性影响(P<0.05),而水流交换率及两者交互作用对该4种物质浓度均无显著性影响(P>0.05)。图3中极端富营养化海水环境下的石莼Car/Chl a和SC/SP比值最大,近岸富营养化海水环境下次之,养虾场废水环境下最小,且SC/SP比值在养虾场废水环境下随水流交换率的增加而增大,且营养盐浓度对两类物质比值均有显著性影响(P<0.05)。
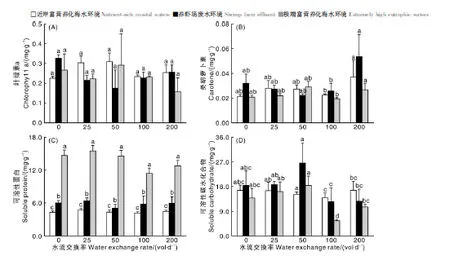
A, 叶绿素a浓度;B, 类胡萝卜素浓度;C, 可溶性蛋白浓度;D, 可溶性碳水化合物浓度。A, Concentrations of Chlorophyll a; B, Concentrations of carotene; C, Concentrations of soluble protein; D, Concentrations of soluble carbohydrate.

表2 应用情境,水流交换率及两者交互作用对羊栖菜生化物质影响的双因素方差分析Table 2 ANOVA analysis of the effect of application scenarios, water exchange rate, and their interaction on biochemical compounds of Sargassum fusiforme
2.3 中试实验
由图5可知,随着流速的增加,羊栖菜的比生长率分别增加80%、14%,石莼则增加41.3%、33.3%。中转速条件下羊栖菜的生物量在前6 d增长缓慢,之后逐渐增加并于第10天达到最大值(6.30±0.29) g。水流交换速率对羊栖菜比生长率无显著性影响(P>0.05)。三转速条件下石莼生物量均生长良好,中转速条件下生物量增长率最大,且从4 d开始生物量一直最大,高转速条件下生物量增长次之。从两者比生长率和生物量数据结合看来,石莼在每个转速条件下的生长都优于羊栖菜,甚至在高浓度条件下石莼的比生长率接近羊栖菜的2倍,但水流交换速率对石莼比生长率无显著性影响(P>0.05)。
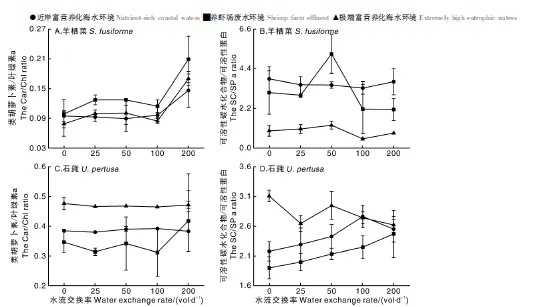
图3 各应用情境下羊栖菜和石莼中类胡萝卜素与叶绿素a比值及可溶性碳水化合物与可溶性蛋白比值Fig.3 Ratios of carotene (Car) to chlorophyll a (Chla) and soluble carbohydrate (SC) to soluble protein (SP) in Sargassum fusiforme and Ulva pertusa under the application scenarios

图4 各应用情境下石莼生化物质浓度Fig.4 Concentrations of biochemical compounds in Ulva pertusa under the application scenarios

表3 应用情境(Ⅰ),水流交换率(Ⅱ)及两者交互作用(Ⅰ×Ⅱ)对石莼生化物质影响的ANOVA分析Table 3 ANOVA analysis of the effect of application scenarios (Ⅰ), water exchange rate (Ⅱ), and their interaction (Ⅰ×Ⅱ) on biochemical compounds of Ulva pertusa

图5 旋转养殖系统示意图及不同水流交换速度下羊栖菜和石莼的比生长率和生物量积累Fig.5 Schematic diagram of rotation cultivation system, SGR and biomass accumulation of Sargassum fusiforme and Ulva pertusa under different water exchange velocity
3 讨论
羊栖菜比生长率受营养盐影响显著,在养虾场废水环境下表现出最大值,且随着水流交换的增加而增大,与Koehl等[35]研究结果一致。可能是由于水流交换率的增加可减少羊栖菜藻体表面的扩散边界层厚度加快二氧化碳和营养盐从介质中向藻体表面的传递速率[21,23],从而促进其藻体生长。在极度富营养化海水应用情境中,羊栖菜藻体叶柄均出现发黑脱落现象,说明高浓度营养盐超过羊栖菜需求的临界值,较低的水流交换率可以缓解高浓度营养盐的毒害作用,但生长明显弱于另外两情境下,过高的水流交换率所带来的水流能量反而加速羊栖菜藻体的脱落[36-38],降低其生长速率。
与羊栖菜相比,石莼在极端富营养化海水应用情境中生长更好,由此表明石莼是海水富营养化修复的良好物种,这与程丽魏、徐永健等[36,39]的研究一致。石莼虽在极端富营养化海水环境下能保持较高的生长率比,但水流交换率达到200 vol·d-1时,藻体出现漂白现象,说明此条件已经不利于石莼生长,导致藻体开始分解。石莼是一种组织C、N、P含量较小的海藻,藻体对C、N、P的贮存量比其他海藻低,因此推测当石莼快速吸收的营养盐远远大于自身合成物质和生长需要时,铵盐在藻体内过度累积造成NH3中毒,导致藻体漂白分解[11]。此外,石莼是由两层细胞构成的叶状藻体,全部藻体细胞暴露在海水环境中[40],水流交换率增大在增加营养盐稳定供给的同时却减少叶面上的扩散边界层,加快营养盐从介质中向藻体表面的传送速率[41],从而加速氨氮的吸收导致NH3中毒,藻体漂白分解。
藻体的光合作用是对外界氮、磷供给最敏感的反应之一[42]。营养盐供给的变化导致羊栖菜中叶绿素a和类胡萝卜素含量略有波动,但无显著差异,这与程丽巍[36]的研究一致。在养虾场废水环境下,Car/Chla比值达到最大,说明在此条件下光合色素中类胡萝卜素发挥主要作用,吸收和传递光能,促进藻体生长。某些藻类中叶绿素a含量与营养盐浓度的关系并不密切,而与其他环境因子如光照、温度存在一定的相关关系[39,43],但营养盐浓度的增加可促进羊栖菜生长。因此,营养盐浓度增加对羊栖菜生长的促进可能并不是通过影响光合色素(叶绿素a和类胡萝卜素),而可能是通过影响其色素—蛋白质复合物合成进而弥补光的吸收与传递[44]。高浓度营养盐增加了羊栖菜藻体可溶性蛋白含量,但无法抵消高浓度营养盐对藻体其他生理生化活动的毒害作用[37]。
石莼Chl a和Car含量随营养盐浓度和水流交换速率的增加呈上升趋势,说明营养盐供给可以促进石莼光合反应中捕光色素类胡萝卜素和叶绿素a含量来增强光合作用,促进藻体生长[39, 45-46]。在低、中浓度营养盐即近岸富营养化海水、养虾场废水环境下,Car/Chla比值相对较低,说明石莼藻体主要合成叶绿素a来提高光合反应,而在高浓度营养盐即极度富营养化海水应用情境下Car/Chla比值较高,反映了叶绿素a合成受阻,藻体则主要通过提高类胡萝卜素含量来弥补藻体吸收光能[47]。SC/SP比值在极度富营养化海水应用情境下达到最高,说明石莼藻体所产生的能量主要用于合成碳水化合物而非蛋白质。而养虾场废水应用情景下,石莼则会将更多能量用来合成蛋白质,且水流交换率的增加可在一定程度上提高其合成速率与含量。碳水化合物含量在高浓度营养盐下增长可能是与光合作用产物合成速率和消耗速率有关[48],石莼藻体的光合作用产物合成速率大于其消耗速率,从而导致可溶性碳水化合物的积累[49]。
但中试实验中,极端富营养化海水环境中藻体并未出现漂白现象。据此推测,水体与石莼的相对运动使由两层细胞构成的石莼藻体叶片大面积堆叠在一起,增加扩散边界层,降低营养盐从介质中向藻体表面的传送速率[41],缓解NH3中毒。因此,自主设计的旋转系统能够有效提高水流交换并减少过量营养盐对藻体带来的损害,而近年来人工上升流技术[50]的不断发展使水流控制成为可能,这也暗示了研究优化大型海藻养殖中水流交换的重要性。
谢辞:感谢浙江海洋水产养殖研究所林少珍工程师、张鹏研究员及洞头基地相关工作人员在藻种和实验场地、器材方面提供的帮助。
猜你喜欢
中国海洋大学学报(自然科学版)(2022年8期)2022-08-15 06:35:20
防护林科技(2020年9期)2020-11-09 01:45:24
中南林业调查规划(2018年3期)2018-11-24 03:18:48
水利技术监督(2017年6期)2017-12-19 13:28:17
科技创新与应用(2017年20期)2017-07-15 20:43:02
广东海洋大学学报(2015年3期)2015-12-22 10:05:28
中南林业调查规划(2015年3期)2015-12-20 05:22:31
应用海洋学学报(2015年3期)2015-11-22 07:39:10
湖泊科学(2015年4期)2015-02-17 06:43:49
应用海洋学学报(2014年2期)2014-11-26 01:20:48
