
Oxygenation in carbonate microbialites and associated facies after the end-Permian mass extinction:Problems and potential solutions
2018-08-22 01:31:42StephenKershawHaoTangYueLiLiGuo
Journal of Palaeogeography 2018年1期
Stephen Kershaw,Hao Tang,Yue Li,Li Guo
aDepartment of Life Sciences,Kingston Lane,Brunel University,Uxbridge UB8 3PH,UK
bState Key Laboratory of Oil and Gas Reservoir Geology and Exploitation,Southwest Petroleum University,Chengdu 610500,Sichuan Province,China
cCAS Key Laboratory of Economic Stratigraphy and Palaeogeography,Nanjing Institute of Geology and Palaeontology,Chinese Academy of Sciences,Nanjing 210008,Jiangsu Province,China
dCASP,Department of Earth Sciences,University of Cambridge,Cambridge,CB3 0UD,UK
Abstract Carbonate sediments deposited in normally-oxygenated shallow ocean waters of the latest Permian period,immediately prior to the end-Permian mass extinction,contain well-developed diverse shelly faunas.After the extinction of these skeletal metazoans,the sediments commonly comprise microbialites(regarded by most authors as benthic)and associated facies bearing evidence interpreted by many authors to indicate reduced oxygenation of the shallow ocean waters.However,the evidence of oxygenation state is inconsistent and the sequences have gaps,indicated in the following 5 points:1)Shelly fossils occur commonly in post-extinction shallow marine limestones,and likely to have been aerated in contact with the atmosphere.Nevertheless,although the largest mass extinction in Earth history may have caused reduced body size in shelly organisms,such reduction is arguably due to environmental stress of lowered oxygenation.Discriminating between these controls remains a challenge.2)Abundant pyrite framboids in many post-extinction limestones are interpreted by several authors as indicating dysoxic contemporaneous waters,so the organisms that lived there,now shelly fossils,were dysaerobic.However,verification is problematic because pyrite framboids scattered amongst shelly fossils cannot have formed amongst living organisms,which need at least some oxygen;synsedimentary framboid formation requires anoxic conditions in the redox boundary where sulphate-reducing processes work.Thus,framboids and shelly fossils found together means taphonomic mixing of sediments,destroying original depositional relationships so that it is not possible to determine whether the shells were aerobic or dysaerobic prior to sediment mixing.Furthermore,diagenetic growth of framboids is possible,as is import of previously-formed framboids from deeper water during upwelling.Thus,there is no proof of an environmental link between framboid size and occurrence,and contemporaneous oxygenation in these post-extinction shallow water facies,so we question the validity of this model in those facies,but consider that the model is valid for deeper water facies.3)Some publications provide evidence of oxygenation,from redox-sensitive elements in post-extinction limestones,while others indicate low oxygen conditions.Redox-sensitive geochemistry requires further work to explore this issue at higher resolution of sampling than has been so far applied.4)Biomarkers recorded in some post-extinction facies contain evidence of anoxic conditions(including green sulphur bacteria)but other examples lack these,which may be indicate fluctuations of water oxygenation.However,a key issue that has not yet been resolved is determination of whether biomarkers were imported into the sites of deposition,for example by upwelling currents,or formed where they are found.Thus,there is currently no verification that biomarkers of low oxygen organisms in shallow water settings actually formed in the places where they are sampled.5)The common occurrence of small erosion surfaces and stylolites represents loss of evidence,and must be accounted for in future studies.The oxygenation state of post-end-Permian extinction shallow marine facies continues to present a challenge of interpretation,and requires high-resolution sampling and careful attention to small-scale changes,as well as loss of rock through pressure solution,as the next step to resolve the issue.
Keywords Stromatolite,Thrombolite,Microbialite,Permian—Triassic boundary,Pyrite,Anoxia
1. Introduction and aims
The state of ocean oxygenation through the end-Permian extinction event is a topic of intense scrutiny in numerous publications and a range of evidence exists using:(a)sedimentary facies and fossils(e.g.,Algeo et al.,2011,2013;Baud et al.,2007;Farabegoli et al.,2007;Forel et al.,2009;Kershaw et al.,2007;Twitchett and Wignall,1996;Xie et al.,2010);(b)biomarkers(e.g.,Grice et al.,2005a;Luo et al.,2013;Nabbefeld et al.,2010a;Xie et al.,2005,2007;Zhou et al.,2017);(c)physical proxy of pyrite framboids(e.g.,Bond and Wignall,2010;He et al.,2013;Liao et al.,2010,2017;Tian et al.,2014);and(d)geochemical proxies of oxygenation(e.g.,Algeo et al.,2007,2008;Collin et al.,2015;Lau et al.,2016;Meyer et al.,2008;Nabbefeld et al.,2010b;Ryskin,2003;Song et al.,2014).Although there is good evidence for reduced oxygen conditions in the deeper ocean system(e.g.,Bond and Wignall,2010;Lau et al.,2016;Wignall and Newton,1998),even in shelf settings(e.g.,Fenton et al.,2007;Tian et al.,2014),there is continuing debate about the degree to which shallow shelf waters were oxygenated,important because these are environments of many post-mass-extinction facies including carbonate microbialites that have become symbolic of post-end-Permian extinction environments.This paper focuses on carbonate microbialites that formed after the extinction of skeletal metazoans(therefore considered in this paper as postextinction microbialites)and their associated facies,because they are very common in shallow marine facies across the end-Permian extinction;however,shallow marine clastic sequences in northern Pangaea(e.g.,Wignall et al.,2016)containing evidence of anoxia indicate the widespread nature of the oxygenation issue.
Because most localities show microbialites cross the Permian—Triassic boundary as defined by the first appearance of the Hindeodus parvus conodont(e.g.,Jiang et al.,2007),we refer to them as Permian—Triassic boundary microbialites(PTBMs,see also Kershaw et al.,2007,2012a).Before extinction there are:high diversity biotas(e.g.,Chen and Benton,2012),rare pyrite framboids(popular indicators of contemporaneous environmental oxygen levels,see Bond and Wignall,2010,demonstrating rare framboids in some sections before the extinction),and limited evidence of the types of biomarkers indicative of low environmental oxygen(e.g.,Luo et al.,2013 show aerobic biomarkers,while Nabbefeld et al.,2010a show fluctuations that indicated changes in oxygenation).After extinction of most skeletal metazoans there are:low diversity biotas(e.g.,Erwin,2006),and abundant widespread microbialites(e.g.,Baud et al.,2007),abundant pyrite framboids in most PTB sites(e.g.,Bond and Wignall,2010);some evidence of biomarkers for extreme anoxic conditions of photic zone euxinia(Grice et al.,2005b;Luo et al.,2013;Nabbefeld et al.,2010a);and redox-sensitive geochemical indicators that the environments for the PTBMs were normally oxygenated(e.g.,Collin et al.,2009).Leiet al.(2017)interpreted fluctuating oxygenation conditions in the earliest Triassic using a combined approach of Fe—S—C—Mo geochemistry through the Late Permian to earliest Triassic in a carbonate ramp,presumably too deep for PTBMs in Hubei,South China.Thus,the evidence for oxygenation of PTBMs from various sources has not yet resolved the redox status of the microbialites and their associated facies.
The aim of this study is to explore contradictions and variations of evidence for oxygenation levels in shallow marine post-extinction carbonate facies after the end-Permian event in an attempt to find resolution of the oxygenation problem.The results show there is continuing uncertainty regarding oxygenation of environments now preserved as carbonate rocks,and this paper tries to provide focus for future research.
2. Background features of PTBMs
This section summarizes the key features of PTBMs prior to description of oxygen-related aspects in Section 3.The information in this section is important for understanding facies relationships of the microbialites of relevance to oxygenation issues.
2.1. Age,duration and occurrence of PTBMs
Across the end-Permian extinction,the change from high to low diversity of shallow marine biotas is well-documented(e.g.,Chenand Benton,2012;Erwin,2006),followed by a recovery of at least 4 million years,to 8—9 million years(Chen and Benton,2012).The extinction,and thus change to low diversity biotas,occurred very quickly,perhaps as short as 160 ka (Erwin,2006).Baresel et al.(2017)attempted to determine the length of time that the PTBMs grew,proposing a period of about 14 ka,but unfortunately their calculated error bar of±57 ka creates difficulty in defining the period of growth with any certainty,because it is of course impossible for rocks to be deposited in a negative number of years.Post-extinction microbialite sequences vary in thickness from the thinnest recorded at 6 cm,to the thickest at about 20 m(Kershaw et al.,2012a)and were facies-controlled.The stratigraphic age of PTBMs ranges from latest Permian conodont biozones(probably Hindeodus praeparvus Zone)but are most represented in the Hindeodus parvus Zone and appear to have been terminated in the early Griesbachian Isarcicella staeschei Conodont Biozone(e.g.,Ezaki et al.,2003,2008;Tang et al.,2017).However,erosion of boundary facies and the paucity of conodonts so far recorded in PTBMs prevent precise determination of the beginning and end of PTBM growth in relation to the established conodont zone system.
PTBMs are largely within the Tethys Ocean and are geographically variable.A general pattern demonstrates that PTBMs in western Tethys are sediment dominated,and those in eastern Tethys(i.e.the South China microcontinent) are calcimicrobedominated(Kershaw et al.,2016).PTBMs are generally recognized to have formed as benthic structures,and it is important to verify this because of the implications for the oxygenation issue.South China PTBMs are selected for illustration because their features show the benthic nature of PTBMs very well,an issue discussed later.Fig.1 shows a representative example of South China calcimicrobes,which constructed a digitate architecture;and which have been shown to contain pyrite framboids discussed later.Fig.2 shows the digitate detailed nature of the structure composed of upward-branching microbial material and Fig.3 illustrates the filled framework of the calcimicrobe showing it was a constructed frame.Fig.4 presents evidence of the nature of the constructing organism,interpreted here as a solid micritic mass that becomes commonly recrystallized,explored later in this paper.Overall,PTBMs indicate short-lived and significant biotic and environmental change after extinction.
2.2. Facies control and post-lithification modifications

Fig.1 Representative cross-sections of calcimicrobe-constructed PTBMs(Permian—Triassic boundary microbialites),from Dongwan site,Huaying Mountains,Sichuan Province in South China.Readers are referred to Kershaw et al.(2007,2012a)for details of PTBMs in other areas.A—Field view showing layers of calcimicrobe structure,with a tabular form in the earlier(lower)part passing upwards into domal structure in the upper part.Layers are separated by stylolites and the form of the base of each layer is not clear.Small red-colored scale in center is 10 cm long;B—Detail of domal part of a PTBM illustrating the digitate nature of the structure,overlain by packstone of shelly micrite.The yellow box represents the stratigraphic level of C,although the sample in C was collected along strike,outside this photograph.Hammer head is 15 cm in horizontal length;C—Polished vertical section from the same horizon as the yellow box in B showing a digitate framework filled with micrite in this sample.See Figs.2 and 3 for more details.
PTBM growth appears to have occurred during a transgression following latest Permian regression in shallow-marine sequences,so that the boundary between pre-and post-extinction facies is commonly seen as a sharp contact in most places,illustrated by several authors(e.g.,Payne et al.,2007).The nature of the contact is problematic in most locations because of pressure solution,but small areas of unaffected rock reveal a true sharp contact,that has been interpreted as due to either erosion or dissolution,the latter either by terrestrial rain water or by ocean acidification(see Kershaw et al.,2012b;Payne et al.,2010;for discussion).In most locations the contact is a planar surface(sometimes modified as a result of bedding-plane slip due to local folding),and the stylolitic contact between pre-extinction limestones and PTBMs is normally very clearly visible in the field.However,in the South China Great Bank of Guizhou(GBG),there is a more complex facies sequence of change,containing at least three erosion events in the latest Permian facies,with irregular erosion surfaces recognizable in field settings(Collin et al.,2009).In Turkey,a single smoothly eroded surface is recognized in C¸ürük Dag(Kershaw et al.,2011).There is some evidence of uplift prior to completion of the pre-extinction facies sequence,with recorded pendent cements in one GBG site,interpreted by Collin et al.(2009)as indicating relative sea-level fall,although this is disputed(see comment and reply Kershaw et al.,2016;Lehrmann et al.,2016).Wu and Fan(2003)and Wu et al.(2006)explored the possible amount of sea-level fall that may be represented by the final erosion surface,and these authors suggest several tens of meters relative sea-level drop,based on diagenetic features of the rocks.Overall,it is clear that the latest Permian preextinction limestones were fully lithified and eroded prior to deposition of the PTBMs.It is also clear that the early parts of some PTBMs were eroded prior to the main growth of the microbialite,as shown in Turkey(Baud et al.,2005;Kershaw et al.,2011),and erosion may have also occurred in Chinese sequences,because some sites show the first deposit is a stromatolite(e.g.,Adachi et al.,2017)overlain by fabrics that look like thrombolites in the field(in China,most are actually composed of calcimicrobes,visible in thin section),while in most sites there is no stromatolite.It may be that the lack of stromatolite in most Chinese sites is due to erosion of the first-formed component of the microbialite,although there is no evidence of such erosion of early parts of PTBMs in sites so far studied by numerous authors.

Fig.2 Details of digitate calcimicrobe-constructed PTBMs demonstrating upward-branching architecture (A, B, D) that is laterally composed of an anastomosing framework (C). A—C are from Dongwan site, Sichuan Province, South China; D is from Laolongdong site, Chongqing, South China.

Fig.3 Thin section views of calcimicrobe-constructed PTBMs from Laolongdong site,Chongqing,South China,demonstrating the structure formed an open framework with geopetal sediment fills in a vertical section(A)and possible microbial micrite coatings in cavities in a transverse section from a different sample(B)with subsequent wackestone fill.These pictures emphasise the framework structure of the microbialite,discussed in the text.
Erosion surfaces indicate loss of deposited rock,but are valuable because they indicate a dynamic environment involving early lithification of carbonate sediments followed by frequent erosion,thereby demonstrating an active shallow marine setting subject to energy changes.Erosion surfaces and stylolites both interfere with the sequence record by removing evidence of environmental change.Although the majority of stylolites are generally limited in both lateral extent and in the vertical loss of rock,their abundance in PTBMs significantly impedes understanding details of the processes of change within the microbialites.For example,in Sichuan and Chongqing in South China,all sites so far studied by all authors revealed a stylolitic contact between the latest Permian pre-extinction limestones and the base of the microbialite.

Fig.4 A—Vertical section of a small portion of calcimicrobe from Baizhuyuan site,Sichuan Province,South China,showing variation of appearance of the structure.In the center-left,the calcimicrobe comprises a solid micritic mass,while on the left edge and right half the micritic structure passes,with continuity of microbial fabric,to areas composed of a sparitic interior and outer micritic portion.The sparitic parts of the calcimicrobe in this photo show the most commonly preserved appearance of these South China calcimicrobes,giving rise to the view of many authors that the microbe has a Renalcis-like architecture.However,this photograph is presented as evidence to support the view of the present authors that the original structure was made of solid micrite and the sparitic parts are a partially altered form.Other samples,not illustrated here,show the microbial branches may be extensively dolomitized while the micritic in fill is not;consequently we infer the microbialite structure was easily altered to produce the misleading Renalcis-like form;B—For comparison,at the same scale,the structure of Renalcis and Izhella(the latter has angles in the internal wall,whereas the former has a smooth internal wall)from Devonian reefs of the Canning Basin,Australia,showing the contrast between Renalcis and the PTBMs.See text for further discussion.
PTBMs are found in shallow water settings of platform carbonates,but are lacking in the deep shelf position of the Meishan GSSP(Global Stratotype Section and Point).Their presence in deeper rampsettings in northern Hungary is questionable(Kershaw et al.,2012a),even these facies may have been also too deep for them.PTBMs occur in shallow facies on seamounts in Panthalassa Ocean,later accreted in western Panthalassa as part of Japan(e.g.,Sano and Nakashima, 1997). PTBMs are associated with micritic sediments and have not been described from higher energy facies such as oolites,although they are interbedded with oolites in some sites(Kershaw et al.,2012a);it seems they were unable to tolerate high energies.Finally,all PTBM sites so far described show that the microbialite unit was sharply terminated,and overlain by a variety of facies types,different from locality to locality.PTBMs are overlain by either oolite,shelly wackestones or clay-rich micrites and some may have been buried by increasing sedimentation in a rising base level of the Early Triassic.Nevertheless,the precise cause of termination of PTBMs has not been established.
3. Evidence of degree of oxygenation of shallow marine waters after the end-Permian extinction
The four principal lines of evidence relating to oxygenation state of carbonate facies after the end-Permian extinction event are described below.
3.1. Shelly faunas
The presence of abundant shelly faunas in shallow marine limestones is normally considered to be evidence of well-oxygenated conditions,because shallow marine waters are well-mixed by wave and current action due to close proximity to turbulence in the overlying atmosphere.It should therefore be self evident that the shallow marine facies of the PTBMs were oxygenated.However,the literature contains examples of situations where abundant low diversity biotas may occur in dysaerobic conditions(e.g.,Wignall and Hallam,1991;also see Brenchley and Harper,1998 for discussion).In the case of PTBMs,evidence from ostracods(e.g.,Forel et al.,2009)is used to interpret normally oxygenated water,and the presence of conodonts suggests oxygenated facies in which the conodont animals lived.In some sites,mollusk faunas indicate rapid recovery from mass extinction(Hautmann et al.,2011),but these faunas are found in limited sites.In most shallow marine sites,the impact of mass extinction created a unique biota containing microbialites that have no equivalent elsewhere in the geological record,with a fauna generally comprising small-size fossils that may have been influenced by low levels of oxygen.Thus,although sites of diverse faunas may be presumed to demonstrate normal oxygenation,in most places verification from the biotas is not completely certain because of proxies for reduced oxygen levels described in the following sections that leave open the possibility of reduced oxygen levels impacting the organisms.
3.2. Pyrite framboids
In ancient rocks,framboids are commonly interpreted as indicating either reduced levels of oxygen(dysoxia)or no oxygen(anoxia to euxinia)in the water column above sedimentary deposits(e.g.,Bond and Wignall,2010).This concept is based on observations by Wilkin et al.(1996)who demonstrated that in cases of dysoxic water in marginal marine settings of salt marshes on the USA east coast,the framboids which form in the sediment below the sediment—water interface range in diameter from approximately 3—33 microns.In contrast,in the Black Sea,where the water is anoxic below the redox boundary,the framboid range is much smaller,approximately 3—13 microns,with the most reliable indicator of anoxia found where framboids are no larger than about 6 microns.There are several published datasets of framboids from the Permian—Triassic boundary facies that have been used by authors to interpret contemporaneous dysoxia(e.g.,Bond and Wignall,2010).A good example of how framboids have been applied is given by Liao et al.(2010)who sampled for pyrite framboids in the PTBM at Laolongdong site,Chongqing,South China and found that pyrite framboids were absent in the preextinction Changhsingian limestones,but were present in the overlying microbialite and the succeeding clay-bearing micrites of the Feixianguan Formation.Framboid sizes there are around 8 microns,considered acceptable for low oxygen waters.Therefore,under the commonly-applied interpretation,such framboids suggest low oxygen formation of the microbialite.Viewing the oxygenation issue in a wider perspective,Paytan et al.(2012)showed that the34S isotope curve for the last 900 million years demonstrates that the lowest point of the curve,indicating the most anoxic state of the oceans was across the Permian—Triassic transition,and the sharp rise following this lowest point was interpreted by Paytan et al.,as being the result of ocean overturn,bringing low oxygen water to the surface.Such work shows that the dysaerobic model has validity for PTBMs,but there is a problem of verification of the model,considered in the discussion section of this paper.
3.3. Redox-sensitive geochemical data
Redox-sensitive chemistry has been applied to the Permian—Triassic extinction facies and the results are variable.Lau et al.(2016)used uranium data to interpret low oxygen levels in shallow waters resulting from upwelling from deeper low oxygen waters,throughout the Late Permianto Early Triassic.However,Collin et al.(2015)found a negative cerium anomaly in four sites in Tethys(Turkey,Hungary,northern South China and southern South China).Mo,U and V measurements in those sites also show normal oxygenation,so overall their data indicate well-oxygenated PTBM facies.Chen et al.(2015,page 187)warned of the danger of diagenetic change in REE,with MREE enrichment and positive Eu anomalies,present in the dataset of Collin et al.(2015),but corroboration of oxygenated conditions is given by Mo,V and U in that study,along with prominent negative Ce anomalies in all sites studied across Tethys.This consistency reinforces confidence in the quality of the data.
3.4. Biomarkers for low oxygen conditions
In deeper environments evidence of low oxygen conditions in latest Permian and Early Triassic are recorded,with examples at the Global Stratotype Section and Point(GSSP)at Meishan,South China(Cao et al.,2009)where Xie et al.(2005,2007)recorded two episodes of low oxygen conditions from biomarker evidence at Meishan.The Hovea-3 core in Perth Basin,Western Australia(Grice et al.,2005b)also shows some fluctuations of oxygenation in deeper waters.However,in shallower conditions the situation seems to be more complex.Luo et al.(2013)demonstrated change from biomarkers showing oxygenated waters in the latest Permian pre-extinction shallow marine facies in South China,to biomarker assemblages indicating reduced oxygenation in the upper water column.In contrast,Zhou et al.(2017)found no aryl isoprenoid(AI)biomarkers of obligate anaerobic green sulphur bacteria(Chlorobiaceae)in shallow marine sequences in a study of a range of facies and different water depths in the PTB facies,also in South China;their results thus do not present any evidence that the water was reduced in oxygen content.Further complication was indicated by Chen et al.(2011)from biomarker evidence of fluctuations in oxygenation in the shallow water PTBM sequence at Chongyang,Hubei Province,South China.The difference between deeper and shallower settings was emphasized by Luo et al.(2014)who found variation in organic carbon isotope results,which they interpreted to indicate low oxygen conditions in deeper settings but oxygenated conditions in shallow waters,in South China.Overall,the data from biomarkers in relation to oxygenation of the PTBMs is complex,with no clear pattern emerging from work so far.A comparison with clastic facies is appropriate here,noting that Wignall et al.(2016)recorded an extended phase of anoxia through the Griesbachian shallow clastic sequence in Spitsbergen,but there were fluctuations higher in the sequence,again demonstrating inconsistency in oxygenation states of shallow marine post-extinction facies.
4. Discussion
4.1. Microbialite construction and nature
Before discussing the oxygenation of the sediments,a key aspect of the PTBMs is their nature and style of growth.Although there is really no doubt that sediment-dominated PTBMs of western Tethys were benthic constructions of stromatolites and thrombolites,two stimulating studies by Wu et al.(2014,2016)proposed that the South China PTBMs were constructed by pelagic cyanobacteria that were deposited on the sea floor to build the microbialites.Figs.1—3 show the architecture of South China PTBMs and we regard these as benthic,principally because of:1)their facies-limited biostromal form of digitate structure(Figs.1 and 2)that has an amalgamated structure when seen in transverse section(Fig.2C);and 2)common occurrence of upward-branching frame construction(Fig.2D)enclosing cavities containing geopetal sediments(Fig.3A)and in some cases micritic cavity-wall coatings(Fig.3B).In our interpretation,it is very difficult,perhaps impossible,to understand how planktonic cyanobacteria would be able to construct upward-branching anastomosing frameworks on the sea floor if they were deposited effectively as a rain of particles onto the sediment surface.Furthermore,Wu et al.(2014)interpreted the sparitic structure of the calcimicrobes as being cement fillings into spaces left by decayed cyanobacterial organic tissue that had been buried in sediment.However,that interpretation is not consistent with a framework structure that contains open spaces and has varied fills(Fig.3).Furthermore,Fig.4 shows variation of appearance of the calcimicrobe from solid micrite to partly sparitic structure.We here interpret this as indicating that the original microbialite structure was made of micrite and thus is not a Renalcis-like microbe.It more closely resembles the Cambrian microbe Tarthinia(see Lee et al.,2014)in both appearance and scale,althoughas far as weare aware,Tarthinia did not construct digitate forms,suggesting that the South China PTBMs may have been constructed mainly from a unique calcimicrobe that may warrant its own name.We propose that its micritic composition was easily altered to give the misleading impression of a Renalcis-like structure(Fig.4B),but this leaves the problem of why the outer part of the solid micrite(Fig.4A)was less susceptible to alteration;perhaps this part was protected from early recrystallization by an organic coating.Other published photographs(e.g.,Kershaw et al.,2012a)shows that in many cases the microbial fabric consists of only sparite,which we interpret as pervasive recrystallization of an originally micritic structure.In many cases,not illustrated here,the sparitic microbe is represented as coarse crystals of dolomite,while the sediment in fill shows either little alteration or is partly to completely altered to fine crystals of dolomite.Whether this structure is an indication that the calcimicrobe was composed originally of high-magnesium calcite(readily dolomitized)or not remains an open question to be further explored.
Overall,in our opinion,the structures illustrated in Figs.1—4 of this paper are sufficient demonstration that the South China PTBMs were constructed as a framework of benthic calcimicrobes,and the following discussion is presented on that basis.Frame construction including cryptic spaces in PTBMs has also been illustrated from elsewhere in the Huaying Mountains(e.g.,Ezaki et al.,2003)and in other areas,such as Guizhou(Adachi et al.,2004;Lehrmann,1999),China.It thus follows that the suggestion of toxic algal blooms proposed by Wu et al.(2014),as related to the extinction,is not supported by evidence in the microbialite structure of the South China PTBMs.
4.2. Oxygenation of the microbialite and associated facies
Discriminating aerobic from dysaerobic biotas on the basis of shell shape(for ostracods)and taxonomic assemblages in rocks a few tens of thousands of years after the Earth's largest mass extinction has no parallels in Earth history,so distinguishing between these in post-end-Permian sediments is problematic because of a lack of verifiable criteria.Therefore,the proxies described in Section 2 of this paper have great importance.It is clear that redox-sensitive elements and biomarkers show varying oxygen levels,requiring more investigation.The issues regarding pyrite framboids are key to this discussion,so the remainder of this section focuses on framboids.
Perusal of the literature on formation of pyrite framboids makes clear that framboids form only in the redox boundary in anoxic conditions(e.g.,Bond and Wignall,2010).However,even dysaerobic metazoans require at least a minimum level of oxygen; fish,for example need a minimum of about 3 ml oxygen per liter of water(e.g.,Ekau et al.,2010)and cannot live in anoxic locations.Work by Danovaro et al.(2010)on metazoans living in anoxic conditions in the Mediterranean Sea,relates to very small organisms(approximately 100—150 microns long)not consistent with the larger sizes of familiar metazoans(see also the debate on whether such organisms were alive when sampled,presented by Mentel et al.(2016)and Danovaro et al.(2016),which shows this issue is not resolved).For comparison,dysaerobic conditions are defined by oxygen levels of 0.1—1.0 ml/l of water(e.g.,Allaby,2008).In cases where shells and framboids occur mixed together,they must have been mixed either later by taphonomic processes(Fig.5),or potentially the framboids were diagenetic.Thus,published applications of pyrite framboids as indicators of oxygenation levels in contemporaneous sedimentary environments require independent verification.It is also worth noting that in places where framboids are absent,this does not necessarily mean the facies were oxygenated because pyrite requires sufficient iron and may be absent in iron-limited settings.

Fig.5 A—A cartoon vertical section showing that shelly benthic biotas living on the sediment surface in shallow waters after the end-Permian extinction may have been located a short distance above the redox boundary,particularly if bioturbation of sediment was limited due to extinction of many burrowing organisms(trace fossils are rare in facies immediately after the extinction).Sea water is blue,sediment is brown.Pyrite framboids(red stars)form in the redox boundary(blue dashed line)by sulphate reduction,as long as sufficient iron,sulphur and organic matter are available;B—Contemporaneous storms or strong currents have the potential to have mixed framboids with shelly remains,producing bioclastic wackestones to grainstones containing framboids.
An example of how close the redox boundary may be to fully oxygenated organisms in Earth surface environments may be found in the modern bivalve Mercenaria mercenaria in salt marshes,Georgia,USA(Clark and Lutz,1980),see Fig.6.Living shells buried in mud are sealed,and water enters the shell only through siphons from oxygenated water above.A thin layer of pyrite encrusting its shell,except for the uppermost part,shows that the shell lives effectively submerged beneath redox boundary,but is an aerobic organism and needs oxygen>5 mg/l(approximately 3.75 ml/l)for unstressed life(Grizzle et al.,2001),similar to modern fish.The implication is that if such shells were found asbioclastic debrisin postextinction facies,then because pyrite cannot form in dysoxic conditions,such deposits would be proof of taphonomic mixing.Within modern tropical microbialites in Qatar,Perri et al.(2017)found pyrite framboids in the deeper part of a living microbial mat(in a hypersaline peritidal setting),a clear indication that the redox boundary can be very close to the sediment surface,so the potential for taxonomic mixing by storms is not in question.
In contrast to the modern examples,an example from the PTB facies,where verification of oxygenation is needed,is given by Liao et al.(2017),in the Dajiang site in Guizhou Province,South China,a PTBM sequence containing thin beds of bioclastic limestones.Within the bioclastic limestone beds,pyrite framboids found mixed with the shells were interpreted as indicating the shelly organisms were dysaerobic biota;this situation is different from the modern M.mercenaria,and the interpretation by Liao et al.(2017)is of course valid,but not verified.Here,it is important to note that although framboids and dysaerobic shelly organisms can grow at the same time in the same general environment,they mustbe in separate microenvironments,without mixing,because framboids form in anoxic water of the redox boundary,while dysaerobic organisms living above the redox boundary need at least some oxygen(see Wignall and Hallam,1991;Wilkin et al.,1996).In contrast,pyrite framboids deposited in deeper water conditions(e.g.,He et al.,2013;Shen et al.,2016)are not questioned in this paper as indicators of oxygenation of contemporary marine water because taphonomic mixing is unlikely to be an issue.However,for shallow waters,we have identified at least four possible mechanisms that could have brought framboids and shelly organisms together in PTBMs,summarized in Fig.7.

Fig.6 A modern bivalve,Mercenaria mercenaria(see Clark and Lutz,1980)lives almost completely buried beneath the redox boundary on modern salt marshes in Georgia,USA.A and B—The large shell with a prominent coat of pyrite encrusting its surface.Note that in this sample some of the pyrite has been rubbed off in handling the shell;C—A detail of the pyrite coating,enlargement from the central part of the shell in B;D—Diagrammatic representation of the mode of life of M.mercenaria,illustrating its growth below the redox boundary(red line),above which there is no pyrite coating,and located close to the sediment—water interface(black line).Siphons draw oxygenated water through the shell,which is an aerobic organism,contrasting the anoxic environment surrounding it.See text for discussion of the implications of this modern bivalve for the PTBMs.
1)Shells grew in fully oxygenated conditions with the redox boundary a short distance below them,close to the sea floor,so that storm action could easily mix the two.Hofmann et al.(2015)demonstrated that the loss of benthic life after the mass extinction included burrowing organisms,so that the mixing of sediment,a normal process on the sea floor during non-extinction times,would be expected to have effectively ceased,or operated at a low level.The senior author of this paper,in approximately 20 years of study,has observed very little evidence of burrows or bioturbation in the PTBMs.Thus,the redox boundary developed in the sediment is likely to have been very shallow in the sediment so that pyrite framboids may have formed very close to the sea floor,at the same time as an oxygen-requiring biota a short distance above.Consequently,strong currents could potentially have easily mixed the sediment,and therefore framboids,with shell material.
2)Shells grew in dysoxic conditions,again with the redox boundary just below the sediment—water interface,allowing easy mixing.In this case the shelly organisms would have been dysaerobic.
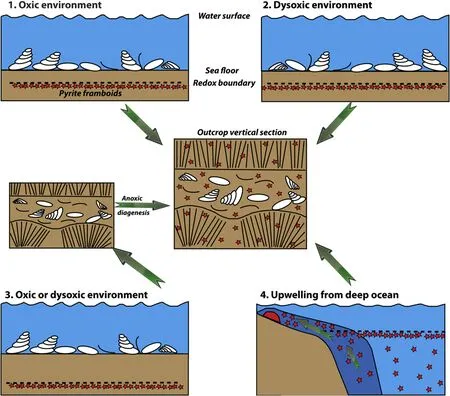
Fig.7 Schematic cross-sections of facies after the end-Permian extinction showing alternative explanations for the presence of shelly fossils mixed in sediment with pyrite framboids,which may lead to the same result,in the center diagram.Blue is sea water;brown is sediment;red stars are pyrite framboids;black dashed line is the redox boundary;red hemisphere in 4 is the microbialite;dark blue area in 4 is upwelled deep water.1)and 2)indicate contemporaneous mixing by current action,in either aerobic or dysaerobic settings respectively;thus discriminating between these oxygen levels is likely not possible.3)indicates early diagenesis to form the framboids;the redox boundary is artificially placed low down in this diagram to emphasis the point that framboids in the central diagram could be derived from diagenesis after the shelly fauna has been affected by turbulence and buried.4)shows the potential for upwelling from deep ocean waters to import framboids,based on observations of suspended framboids below the redox boundary in the modern Black Sea(see Wilkin et al.,1996).The four alternatives are discussed in the text.
3)Pyrite framboids grew diagenetically in the bioclastic limestone.There is no certainty about pyrite framboid size in relation to oxygenation levels because pyrite framboids begin as small crystal aggregates and grow,and framboid abundance depends on availability of iron,sulphur and organic matter.Munnecke and Samtleben(1996)demonstrated very small pyrite framboids(3 micron diameters,which is within the size range used to indicate anoxic conditions in the Black Sea,see Wilkin et al.,1996)in middle Silurian bioclastic limestones associated with reef facies in Gotland,Sweden,richly fossiliferous,in fully oxygenated carbonate facies lacking the types of fossils or sediments that could indicate reduced oxygenation.These framboids were certainly not formed in the same conditions as the facies in which they are found and were either diagenetic or formed below the contemporaneous redox boundary without any oxygenation implications for the biota above.Furthermore,Gotland at that time was in a location a long way from any deep ocean facies(see next point below).
4)The possibility of importing of pyrite framboids by upwelling from deep ocean anoxic settings adjacent to carbonate shelves cannot be ruled out.Kershaw et al.(2016)presented a model by which framboids could be carried up into shelf environments by upwelling,based on the current behavior of Black Sea waters,because of instability of:a)the redox zone(Ostrovskii and Zatsepin,2011,2016);and b)the observed occurrence of framboids suspended in organic matter and therefore not sinking to the sea floor(see Wilkin et al.,1996 for discussion),in contrast to the traditionally viewed character of the high density of pyrite causing rapid export to the sea bed(Tuzhilkin,2008).Kershaw et al.'s(2016)model probably would not work in this mode,because framboids uplifted into the modern oxygenated part of Black Sea waters are likely to oxidize,but if an anoxic water mass upwelled,and invaded shallow facies as proposed for the Permian—Triassic interval by several authors,it is more acceptable.There is a lot of published data supporting interpretations of upwelling in this transition,for example the Sverdrup Basin in northern Pangaea(e.g.,Knies et al.,2013)and the South China Block(e.g.,Shen et al.,2014).
Framboids should be absent from the latest Permian pre-extinction limestones,which may mean diagenetic framboid formation is less likely.Thus the occurrence of framboids in some places suggests diagenetic formation(see Bond and Wignall,2010,showing framboids in some pre-extinction limestones).Note that erosion of pre-extinction limestones recorded in several places indicates that limestone cementation occurred well-developed before the post-extinction facies were deposited,reducing the potential of pyrite to form diagenetically in preextinction limestones,which may partly explain why framboids are uncommon before the extinction.Furthermore,pyrite requires both iron and sulphide(and sufficient organic matter)to be present in the system,and their availability may have differed preand post-extinction.In fact,no framboids in C¸ürük Dag have been found in the microbialite deposit in Turkey(David Bond,personal communication,August 2017),and if the sediments were iron-limited,for example,low oxygen conditions were possible without framboid formation.
Lau et al.(2016)used U evidence to demonstrate anoxia in shallow marine limestones after extinction,which is at variance with other geochemistry data referred to earlier;but this was a broad-scale study(Kimberly Lau,personal communication,May 2017)and small-scale fluctuations of oxygenation are permitted in their work.Upwelling from the Oxygen Minimum Zone(OMZ)may not bring saturated water to the shallows,but could it bring pyrite?If pyrite is not up welled,this emphasizes storm mixing from redox boundary and diagenesis as potential explanations of pyrite framboids in microbialite sequences.
In summary,the current state of knowledge of redox-sensitive geochemistry after the end-Permian extinction contains conflicts of evidence,and likely to be resolved only by high-resolution studies,to attempt to detect fluctuations in oxygenation that may have occurred on a small scale.
5. Conclusions
1)Evidence for oxygenation of shallow marine facies after the end-Permian extinction remains unresolved.Some geochemical data and the presence of shelly faunas in shallow marine carbonates seemingly support a fully oxygenated environment.However,the occurrence of pyrite framboids mixed with microbial structure,and in shell beds interbedded with microbialite,have been used as evidence for dysaerobic conditions in at least some beds.Nevertheless,taphonomic mixing of sedimentary signals remains an issue,because pyrite framboids form in anoxic conditions so could not form in the same micro-environment as shelly faunas which require at least some oxygen,hence interpretations of dysoxia require separate verification.However,the reliability of the pyrite framboid model in deeper waters is not questioned here.
2)Biomarkers for low oxygen conditions in microbialites are equivocal,indicating low oxygen conditions in some sites and normal oxygenation in others.There is also the problem of potential import of biomarkers from ocean circulation(including upwelling),so that whether biomarkers formed in the places where they are found,or not,requires confirmation.
3)Overall,therefore,there is continuing uncertainty about the oxygenation of shallow water facies after the end-Permian extinction,and it is important that future work pays attention to high-resolution study;thus detailed logging is essential,and researchers need to take careful account of the presence of numerous erosion surfaces within microbialites and associated facies.Finally,the importance of the effect of stylolites,which represent potentially significant loss of limestone rock mass,must be considered in interpretations of facies changes in these thin sequences.
Acknowledgements
Steve Kershaw thanks Brunel University for providing facilities for this research,and thanks Ron West,emeritus professor of Kansas State University,for providing the shell of Mercenaria mercenaria used in this study.Hao Tang thanks his PhD supervisor,Prof Xiu-Cheng Tan,for support in Southwest Petroleum University;and acknowledges funding for his work,and academic visits to UK in 2016 and 2017,facilitated by:1)National Scientific and Technology Major Project(Grant No.2016ZX05004002-001),and 2)National Natural Science Foundation of China (Grant No.41602166).David Bond is thanked for helpful discussion of some of the content of this work.
 Journal of Palaeogeography2018年1期
Journal of Palaeogeography2018年1期
- Journal of Palaeogeography的其它文章
- Deeply concealed half-graben at the SW margin of the East European Craton(SE Poland)—Evidence for Neoproterozoic rifting prior to the break-up of Rodinia
- Geologic-seismic models,prediction of shallow-water lacustrine delta sandbody and hydrocarbon potential in the Late Miocene,Huanghekou Sag,Bohai Bay Basin,northern China
- A redescription of the ichnospecies Koreanaornis anhuiensis(Aves)from the Lower Cretaceous Qiuzhuang Formation at Mingguang city,Anhui Province,China
- Evolution of sedimentary facies and fossil communities in the Middle Permian Maokou Formation in Zigui County,Hubei Province,South China
- Trace fossils of an amalgamated storm-bed succession from the Jurassic of the Kachchh Basin,India:The significance of time-averaging in ichnology
- Late Triassic sauropodomorph and Middle Jurassic theropod tracks from the Xichang Basin,Sichuan Province,southwestern China:First report of the ichnogenus Carmelopodus