
Trace fossils of an amalgamated storm-bed succession from the Jurassic of the Kachchh Basin,India:The significance of time-averaging in ichnology
2018-08-22 01:31FranzrsihAlfreUhmanMatthiasAlertiDhirenraPaney
Journal of Palaeogeography 2018年1期
Franz T.Fürsih,Alfre Uhman,Matthias Alerti,Dhirenra K.Paney
aGeoZentrum Nordbayern,Fachgruppe PaläoUmwelt,Friedrich-Alexander-Universität Erlangen-Nürnberg,Loewenichstraße 28,91054 Erlangen,Germany
bInstitute of Geological Sciences,Jagiellonian University,Gronostajowa 3a,30-387 Krakéow,Poland
cInstitut für Geowissenschaften,Christian-Albrechts-Universität zu Kiel,Ludewig-Meyn-Straße 10,24118 Kiel,Germany
dEarth and Planetary Sciences Group/Palaeontology,Manipal Center for Natural Sciences,Manipal University,Manipal 576 104,Karnataka,India
Abstract The uppermost part of the Upper Bathonian Sponge Limestone member,Patcham Formation,of the Jhura Dome of Kachchh Mainland is a thickening-and shallowing-upward succession topped by medium-to thick-bedded hummocky cross-stratified grainstones deposited by storm waves.Occasionally,thin,commonly lenticular,intraclastic—bioclastic silty marl intercalations between the grainstones are highly bioturbated,in contrast to the grainstones,in which,for the most part,trace fossils occur scattered.Large exposures of bedding planes of the grainstones allow the detailed investigation of ichnological features,whereas the high density of traces in the soft marls precludes the identification of any ichnotaxa.Eighteen ichnotaxa have been recorded including Ophiomorpha,Thalassinoides,Taenidium,Gyrophyllites,Chondrites,Dactyloidites,Teichichnus,Bolonia,and Ancorichnus.Except for Ophiomorpha nodosa and Thalassinoides,which generally indicate moderate to high energy conditions and are the dwelling burrows of suspension-feeding to omnivorous crustaceans,the ichnotaxa represent a deposit-feeding behaviour of their producers and thus are characteristic of low-energy environments.The trace fossils form three ichnoassemblages characterized by(1)Ophiomorpha nodosa and Thalassinoides suevicus,(2)?Thalassinoides isp.A,Taenidium,and Bolonia lata,and(3)Ancorichnus.The dominance of traces of deposit-feeders in rocks indicative of high-energy events is counterintuitive and points to their non-contemporaneity.The sediments were deposited during brief high energy events,whereas the trace fossils were produced when,after waning of storms low-energy conditions prevailed.This time-averaging is particularly pronounced in trace fossils that extend vertically downwards and may reach strata deposited under distinctly different conditions. Thus, environmental interpretations based on trace fossils should refer to colonisation surfaces rather than to the sediment surrounding the trace fossils.In the latter case,interpretations may be erroneous,especially when erosion subsequently destroyed sedimentary evidence of their original environment.
Keywords Trace fossils,Middle Jurassic,Kachchh Basin,Taxonomy,Time averaging,Ichnology
1. Introduction
One great advantage trace fossils have over body fossils is that they are,with a few exceptions,preserved in situ and thus appear to be intimately linked to the substrate in which they occur.As such,they are excellent environmental indicators and have been used in numerous cases to support sedimentological interpretations of ancient environments,from the deep sea to coastal and from marine to terrestrial settings(e.g.,Knaust and Bromley,2012).Their use as environmental indicators is,however,impaired by the fact that many trace fossils extend from the surface,at which the producer lived,downwards into sediments that are slightly-to-considerably older.In some cases(e.g.,Ophiomorpha),these three-dimensional structures may extend vertically for several meters.This does not pose a major problem when such trace fossils occur in uniform sediments that record relatively stable environmental conditions.In marginal marine environments,however,where the position of the coastline shifts repeatedly,deep burrows may extend to,and become preserved at levels that record an environment and time distinctly different from the one when the trace maker was alive.A classic example is Ophiomorpha that,produced in nearshore environments,extends downwards into fluvial sandstones(e.g.,Kennedy and Sellwood,1970).Commonly,such non-contemporaneity between trace fossils and sediments occurs at discontinuity surfaces that record erosion and/or changes in relative sea level(e.g.,the marginally marine burrow Psilonichnus penetrating flood plain deposits;Fürsich,1981: fig.3;see also Taylor and Gawthorpe,1993).
Less spectacular,but important for evaluating the significance of trace fossils for the interpretation of the hydrodynamic regime are cases where trace fossils that are generally regarded as characterizing low-energy environments are found in rocks whose lithology(e.g.,grainstones,rudstones)and sedimentary structures(e.g.,large-scale cross-stratification)point to deposition under a high-energy regime.An excellent example illustrating the latter aspect is the succession of hummocky-cross-stratified grainstones at the top of the Upper Bathonian Patcham Formation of the Kachchh Basin,western India.It exhibits a rich trace fossil inventory consisting of both high-and low-energy ichnotaxa,whereby the latter dominate,and thus illustrates distinct time-averaging of ichnotaxa that lived during different hydrodynamic regimes.
2. Geological background
The Kachchh Basin is a small E—W-oriented Mesozoic rift basin at the western margin of the Indian craton(Fig.1)opening towards the west into the then incipient Indian Ocean(e.g.,Biswas,1987,2016).Initiated in Late Triassic times,the sea flooded the basin in the early Middle Jurassic,possibly even during the late Early Jurassic(e.g.,Rai and Jain,2013;see Fürsich et al.,2013 for a summary).In the course of the Jurassic,a thick(>1500 m)pile of marine syn-rift sediments accumulated.In the Early Cretaceous,the basin was in filled and non-marine sedimentation prevailed.During Bathonian times,a hot and partly arid climate promoted the deposition of carbonate sediments.At the beginning of the Callovian,a change to cooler and more humid conditions led to deposition of siliciclastic sediments(Fürsich et al.,2005).During the Late Bathonian, fine-grained carbonate wackestone—marlstone alternations,the so-called Sponge Limestone member(Fig.2),characterized offshore areas and locally contain a rich and diverse fauna of sponges(Mehl and Fürsich,1997).These low-energy background sediments were repeatedly punctuated by several-cm-to decimeter-thick grainstone beds which have been interpreted as the products of storminduced currents(Fürsich et al.,2013: figs.97—98).The member is best exposed in the Jhura Dome(Fig.1)and in the Jumara Dome 38 km further northwest.At the latter locality the thickness and density of event beds increase towards the top of the member,and the last few meters constitute well-cemented flagstones,which are composed of amalgamated hummocky cross-stratified(HCS)grainstones with only rare,thin intercalations of intraclastic silty marl.This thickening and coarsening-upward succession clearly represents shallowing from low-energy environments below the storm wave-base but subjected to frequent storminduced flows to environments within the reach of storm waves.This regressive trend is followed by renewed transgression as evidenced by the basal Callovian Chari Formation(Fig.2),which consists of bioclastic marly silt with intercalations of increasingly thinner and wider-spaced storm- flow beds(see below).
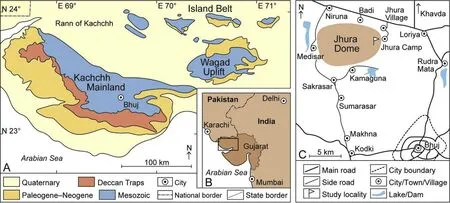
Fig.1 A—Simplified geological map of the Kachchh Basin;B-Position of Kachchh in India(rectangle);C—Locality map.The outcrop investigated is marked by a flag pole(modified after Fürsich et al.,2013).
3. The section
A complete section of the Sponge Limestone member is exposed in a valley east of the highest peak of the Jhura Dome (co-ordinates: N23°24′29′′,E69°35′45′′;Fig.1).The top 1.2 m of the Sponge Limestone member are well exposed at the base of a steep hill slope in the eastern part of the dome(coordinates: N23°24′33.2′′, E69°35′53.9′′and N23°24′32.9′′,E69°35′52.8′′),presenting both cross sectional views and large bedding planes,most of the latter covering an area of 10—20 m2(Fig.2).Altogether 15 beds have been investigated in detail.They strongly vary in thickness and commonly exhibit a lenticular shape(Fig.3).Two lithologies are represented.The dominant lithology is a well-sorted, finegrained bio-intra-grainstone (Fig. 4). Beds are 2—27 cm thick,and within individual beds the thickness varies strongly(e.g.,between 2 cm and 27 cm in the case of bed 14;Fig.5).The components consist of sub-rounded micritic intraclasts,benthic foraminifera,echinoderm fragments,and shell fragments.Many of the foraminifera and intraclasts are stained black,clear evidence that they have been reworked from older beds(Fig.4).The lowermost bed exhibits numerous flattened intraformational pebbles,most of them between 2 cm and 3 cm in diameter,which consist of dark-grey intraclastic marl.
The second lithology is a dark-grey bio-and intraclastic marl. Within the investigated succession, four of these marl beds occur intercalated between grainstone beds.They range from 2 cm to 32 cm in thickness.They all are strongly lenticular relicts filling depressions of the underlying grainstone and commonly disappearing laterally towards the top of the hummocks(Fig.5).
The two lithologies differ in their degree of bioturbation.The marl beds are heavily bioturbated,but the high trace fossil density,compaction,and soft nature make it impossible to identify individual ichnotaxa.In the grainstones,in contrast,except for small patches on their top,the burrow density is low,so that individual burrows are easily discerned.Crossing over of traces is rare.

Fig.2 Stratigraphic position of the investigated rock unit(top of the Upper Bathonian Sponge Limestone member)(left)and large bedding planes of the amalgamated grainstones(storm beds)in the southeastern part of the Jhura Dome.The boundary to the overlying Callovian Chari Formation,a predominantly fine-grained siliciclastic unit,is stippled.Note that the basal Chari Formation still contains thin pack-and grainstone beds alternating with intraclastic marly silt layers before argillaceous to marly silts dominate.Siliciclastic grain size units:cl=clay;si=siltstone;fs= fine sandstone;ms=medium sandstone;cs=coarse sandstone;g=gravel;p=pebble.Carbonate classification:m=mudstone;w/f=wackestone/ floatstone;p=packstone;g=grainstone;r=rudstone.
4. Trace fossil inventory
The following ichnotaxa all occur on the upper surfaces of the grainstones and are therefore either positive or negative epireliefs.In addition,some traces are preserved as full reliefs.Altogether 18 ichnotaxa have been recorded,some of them in open nomenclature.In the following,they are briefly characterized with respect to their morphology,ethology,and trophic regime(see also Table 1).
Ancorichnus isp.(Fig.6A and B)
This ichnotaxon occurs in full relief at the top of grainstone beds 10 and 12.It is a non-branched,cylindrical,horizontal burrow with a diameter of 0.2—0.3 cm and 0.3—0.4 cm,respectively.The fill is identical to the surrounding matrix.The course of the burrows is irregularly sinusoidal with a variable wave length(commonly around 4—5 cm)and low amplitude.The burrows exhibit thin,dense meniscate back fills and were lined by a material softer than the burrow fill,which has now weathered away.In bed 12 the ichnotaxon occurs in great abundance in the troughs of current ripples and follows the direction of the troughs.
The burrows resemble Ancorichnus Heinberg,1974,which is characterized by meniscate filling and a mantle.The trace fossil described as Muensteria from the Middle Jurassic of Greenland (Heinberg and Birkelund, 1984: fig. 10B) and included in Ancorichnus Heinberg,1974 by Keighley and Pickerill(1994)is very similar but less winding.
Ancorichnus is a pascichnion produced by a depositfeeding organism.This interpretation is corroborated by the preferred position of the traces in ripple troughs where particulate organic matter preferentially accumulates.
?Bergaueria isp.
On one bedding plane several circular negative epireliefs with sub-vertical margins and a diameter of 5—6 cm are found.The depth of the depressions,which are filled with marly material,is up to 2.5 cm.

Fig.3 Details of the amalgamated storm beds.Hammer length 33 cm for scale.A—The hummocky cross-stratified(HCS)grainstones(lower half of the photograph)form the top of the Upper Bathonian Patcham Formation and the top of a thickening-and coarsening-upward succession.This up-shallowing succession is followed by bioturbated packstones of the basal Chari Formation(Lower Callovian),which indicate renewed deepening(upper half of photograph);B—Current-rippled top of a HCS grainstone bed overlain by the next HCS unit;C—Large scour at top of a HCS grainstone bed.
Too few details are available to firmly place the trace fossils in an ichnotaxon.They may correspond to Bergaueria,which is generally interpreted as a resting trace(cubichnion)of organisms such as sea anemones(e.g.,Alpert,1973;Pemberton et al.,1988).Most likely,the producer has been a suspension-feeder.

Fig.4 Thin-section of hummocky cross-stratified grainstone,composed chiefly of bioclasts(benthic foraminifera and echinoderm and bivalve fragments)and intraclasts.Dark-stained foraminifera and intraclasts are reworked from older strata.
Bolonialata Meunier,1886(Fig.6C—E)
The slightly positive epireliefs of this trace fossil occur on three bedding planes as a horizontal,straight,non-branched,bilobed ribbon which tapers towards one or both ends.The ribbon is composed of biserial transverse pads separated by a median furrow.The faintly to distinctly arched pads are inclined with respect to the bedding plane and are manifested on the surface as arcuate ridges.The width of the ribbon varies between 1.8 cm and 4 cm,the thickness of individual pads between 0.2 cm and 0.5 cm.The greatest observed extension of the ribbon was 8 cm.
The ribbons closely correspond to Bolonia lata as recently described and figured by Schlirf(2000).The trace fossil has presumably been produced by the lateral movement of an infaunal deposit-feeder through the sediment and thus corresponds to a pascichnion.
Chondrites targionii(Brongniart,1828)(Fig.6F)
Chondrites targionii is restricted to two levels within the succession where it occurs as horizontal full reliefs on the upper bedding surfaces.The tunnels are 0.1—0.12 cm wide,occasionally slightly curved,and are filled with marl that is slightly darker than the matrix.Dichotomous branching is the rule,but not exclusively so.Phobotactic behaviour(avoiding contact with earlier produced burrow segments)is generally developed.The burrow systems closely resemble those figured by Fu(1991:pl.1, figs.A,C,D;see also Uchman,1999;Uchman et al.,2012).
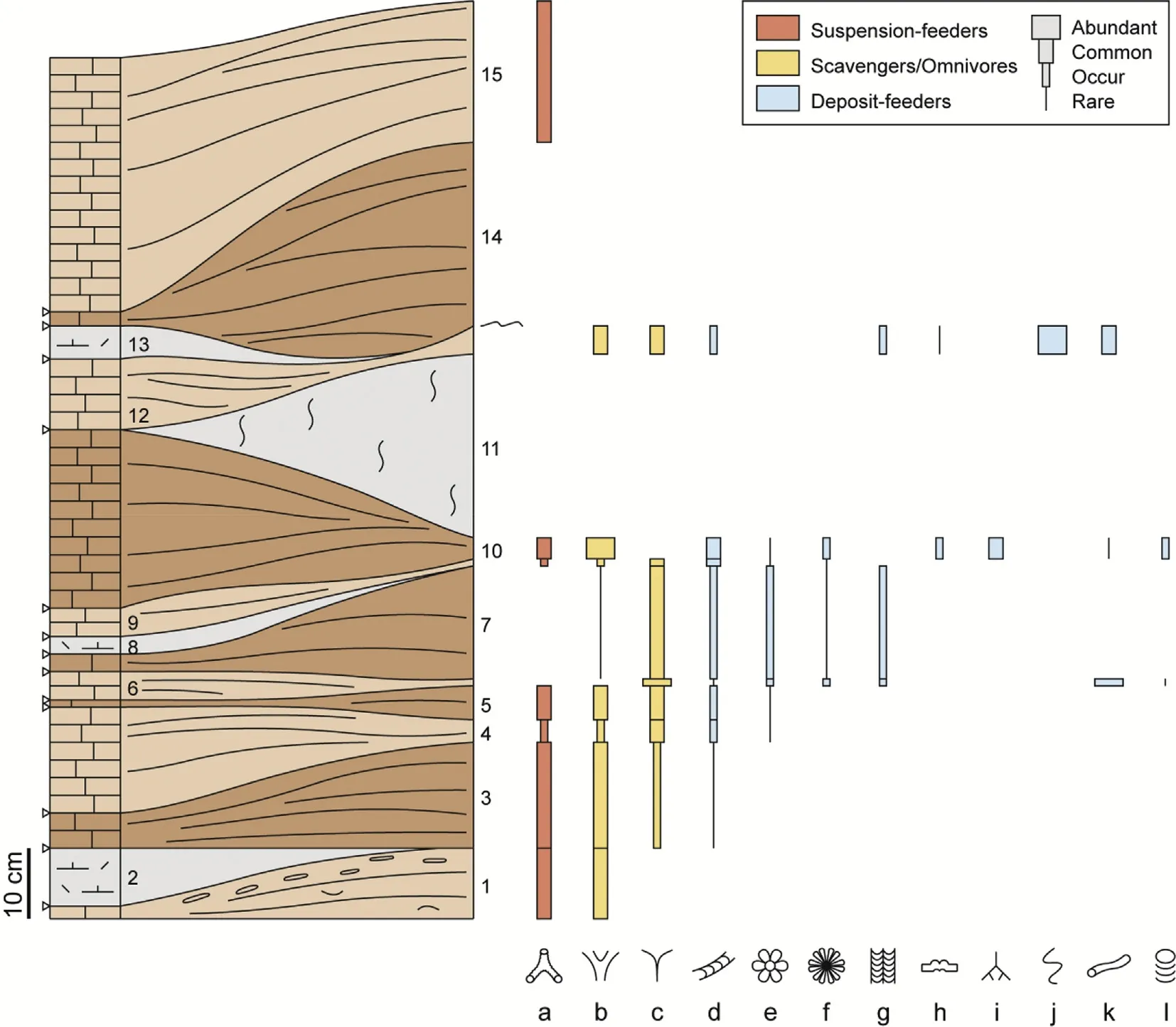
Fig.5 Schematic section through the top 130 cm of the Sponge Limestone member showing the amalgamated nature of the hummocky crossstratified grainstones,the lenticular nature of the intercalated intraclastic—bioclastic marl,and the distribution of the characteristic trace fossils.1—15=bed numbers;a=Ophiomorpha nodosa and O.irregularis;b=Thalassinoides suevicus;c=?T.isp.A;d=Taenidium cf.diesingi and T.serpentinum;e=Gyrophyllites geryonides;f=Dactyloidites ottoi;g=Bolonia lata;h=Gyrochorte comosa;I=Chondrites targionii;j=Ancorichnus isp.;k=Palaeophycus isp.;l=?Teichichnus rectus and T.cf.patens.For key of symbols see Fig.2.
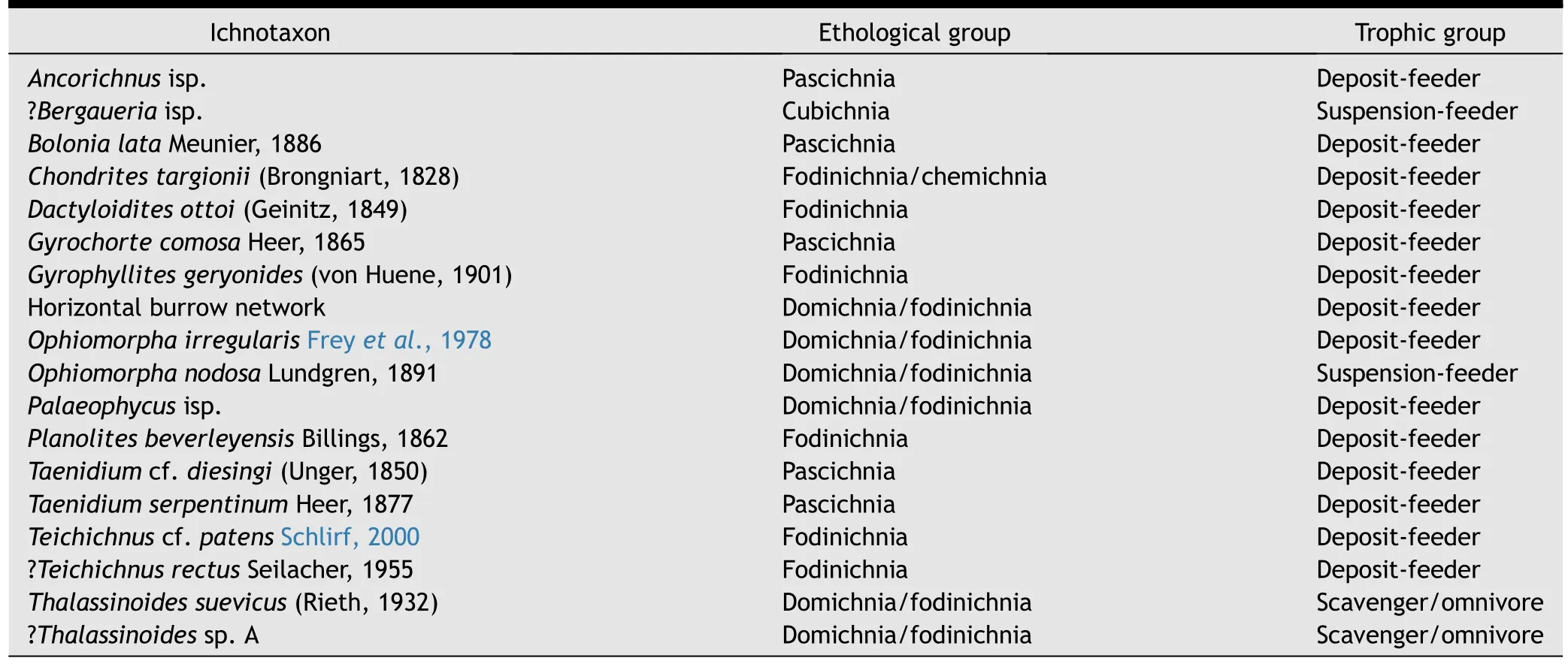
Table 1 List of ichnotaxa occurring in the hummocky cross-stratified grainstones of the top of the Sponge Limestone member,SW Jhura Dome.

Fig.6 Trace fossils from the upper surfaces of amalgamated grainstone beds.A—B—Ancorichnus isp.;C—E—Bolonia lata Meunier,1886;F—Chondrites targionii(Brongniart,1828);G-I—Dactyloidites ottoi(Geinitz,1849).
Chondrites has been interpreted as a fodinichnion or as a chemichnion(e.g.Bromley,1996;Chen et al.,2011,2015;Fu,1991).
Dactyloidites ottoi(Geinitz,1849)(Fig.6G—I)
A stellate structure,4—5 cm in diameter,preserved as full relief or as negative epirelief.Its center is occupied by a cylindrical,vertical shaft which is 0.3—0.4 cm in diameter.The probably originally cylindrical primary radial rays,now compressed by compaction,number around 16 but are difficult to count because they branch repeatedly so that at the external diameter the number of rays may be as high as 40.These secondary radial elements appear to branch off only on one side of the primary rays(cf.Fürsich and Bromley,1985;Wilmsen and Niebuhr,2014).In this way,the sediment around the central shaft could be totally reworked by the producer.There are no sign of several tiers as in the example from the Cretaceous of West Greenland(Fürsich and Bromley,1985).The existence of a downward-or upward-directed spreite could not be verified due to the preservational mode.The fill of the rays displays,however,very indistinct arcuate laminae as in the West Greenland example.Fig.6G shows a cluster of five closely adjacent specimens,most of them circular,but one only fan-shaped with radial elements just forming a half circle.This is most likely a result of the fact that the neighbouring space had been already bioturbated by an adjacent organism.
Recently,Belaústegui et al.(2015)argued that D.ottoishould be transferred to the ichnogenus Haentzschelinia Vyalov,1964,a view already adopted by Häntzschel(1970,1975)and Vyalov(1989).According to the last authors,diagnostic differences are the larger size of Dactyloidites,its Cambrian age,and the more numerous radial elements of Haentzschelinia.Schweigert(1998)argued that Haentzschelinia is mainly preserved as half circle and not fully radial.In the present material,however,most specimens are circular,incomplete circles being rare and explained as due to space limitation(see above).In conclusion,the arguments for separating Dactyloidites and Haentzschelinia at the ichnogeneric level are not stringent and the ichnospecies ottoi should therefore remain in Dactyloidites,which has priority,at least until the type Cambrian material will be better understood.
The burrow pattern and the evidence of avoiding already bioturbated areas clearly support the interpretation of D.ottoi as the burrow of a deposit-feeding organism which exploited the sediment in a very systematic manner.
Gyrochorte comosa Heer,1865
Gyrochorte is a rare trace fossil on the investigated bedding planes,found only on two of them and is seen only for 4 cm.The width of the positive epirelief,which consists of an elevated ribbon divided by a furrow in the middle,is 0.3 cm,in a second example 0.8 cm.Due to their poor preservation,the traces do not exhibit the characteristic plait pattern.
Gyrochorte is the trace of a deposit-feeding wormlike organism,which moved through the sediment in an oblique position(e.g.,Fürsich et al.,2017)and can be termed a pascichnion.It is commonly associated with thin ripple-bedded sandstones and siltstones that alternate with finer-grained sediments,although it also occurs occasionally in calcareous mudstone(e.g.,Chen et al.,2011).As such its preservation is tied to closely adjacent internal surfaces.
Gyrophyllites geryonides(von Huene,1901)(Fig.7A and B)
Gyrophyllites geryonides occurs as a negative epirelief.Around a central vertical shaft,0.5 cm in diameter,12 unbranched petal-shaped lobes are radially arranged.The diameter of the circular structure is 5—6 cm.The preservation of the radial lobes as negative epirelief suggests that they were filled with softer material.
Gyrophyllites has been interpreted as fodinichnion(e.g.,Fürsich and Kennedy,1975;Fu,1991)of a wormlike organism such as a polychaete or echiuran worm(Strzeboénski and Uchman,2015).Overhanging walls of the negative epireliefs testify that the producer,dwelling in the vertical burrow,has not been feeding at the sediment—water interface.Instead,it burrowed from the top and mined the sediment in a horizontal radial fashion.As is so often the case,the best preservation is at internal sedimentary interfaces.Fürsich and Kennedy(1975)demonstrated that in some Gyrophyllites the radial structures are vertically stacked at several levels within the sediment,separated by undisturbed sediment and connected by the vertical shaft.As only bedding planes were available for observation,this could not be verified in the present material.
Strzeboénski and Uchman(2015)recently described Gyrophyllites kwassizensis Glocker,1841 from Upper Cretaceous-Paleocene rocks of the Carpathians.Morphologically,their material is very close to G.geryonides.Glocker's ichnospecies would have priority,but according to Strzeboénski and Uchman(2015)the lobes of kwassizensis were produced and filled with sediment from below and not from above as in the present case.This behavioural difference expressed in morphological details is significant enough to justify their separation at the ichnospecies level.
Horizontal burrow network(Figs.7C and 8).
A rare network of tiny horizontal cylindrical burrows(diameter 0.1—0.2 cm)is preserved as full relief on a bedding plane.The moderate to poor preservation of the traces and the common crossing of burrows makes it difficult to recognize a characteristic pattern.However,Y-to T-shaped branching seems to occur,which would suggest that the producers might have been juvenile crustaceans.Whether the burrows formed a true network of connected tunnels could not be established with certainty,but appears likely.
In ethological terms,the burrow network is a domichnion;the mode of feeding of its inhabitants is,however,not clear.
Ophiomorpha irregular is Freyet al.,1978(Fig.7E and F)

Fig.7 Trace fossils from the upper surfaces of amalgamated grainstone beds.A-B—Gyrophyllites geryonides(von Huene,1901);C—Horizontal burrow network;D—Palaeophycus isp.;E-F—Ophiomorpha irregularis Frey et al.,1978;G-H—Ophiomorpha nodosa Lundgren,1891.
The ichnospecies was originally named irregulaire by Frey et al.(1978).This name contains two mistakes:a spelling mistake(the second “i” is superfluous as the Latin adjective is irregularis)and the ending,which has to be “-is” as Ophiomorpha is“female”.The name of the ichnospecies is therefore herewith changed to Ophiomorpha irregularis which is,according to the rules of the International Code of Zoological Nomenclature(ICZN),a nomen correctum.
Diagnostic features of Ophiomorpha irregularis are the horizontal meander maze,the scarcity of vertical elements,and a wall that consists of irregular,tapering pellets(commonly forming“ flame structures”;Fig.7F)(Bromley and Pedersen,2008;Leaman et al.,2015).In some cases the pelletal lining has been mineralised by iron hydroxide and is therefore more strongly cemented than the burrow fill,which is identical with the surrounding grainstone matrix.As can be seen in Fig.7F,the distribution of pellets can be quite uneven.The cylindrical burrows are 0.5—0.7 cm in diameter and the lining is 0.2—0.3 cm thick so that the total diameter ranges from 1.0 cm to 1.3 cm.Back fill structures are rarely seen.

Fig.8 Sketch of horizontal burrow network.
Like O.nodosa,O.irregularis has been interpreted as the product of decapod crustaceans,but in contrast to the former ichnotaxon its producers probably were deposit-feeders(Leaman et al.,2015).
Ophiomorpha nodosa Lundgren,1891(Fig.7G and H)
Ophiomorpha nodosa occurs as mainly horizontal cylindrical burrows preserved as negative epirelief or full relief,1—1.5 cm,rarely up to 2.5 cm(bed 5)in diameter,with common Y-shaped branchings.Its characteristic feature,i.e.the pelleted burrow wall is,however,often poorly developed(Fig.7H).Vertical elements are rare.
O.nodosa is generally interpreted as a domichnion of suspension-feeding decapod crustaceans such as callianassids(e.g.,Frey et al.,1978;Weimer and Hoyt,1964).
Palaeophycus isp(Fig.7D)
Palaeophycus isp.is a long,horizontal,straight to slightly curved,unbranched,cylindrical burrow,0.4 cm in diameter,with a distinct lining that consists of a slightly darker marly material.The fill is identical to the surrounding matrix.The burrow could be traced for up to 44 cm without showing any sign of branching.
The comparatively thick lining suggests that the burrows were kept as open tunnels and served as domichnia and possibly for feeding(category fodinichnia).The feeding mode of the producer is more difficult to interpret:the producers may have been scavengers or selective deposit-feeders.A suspension feeding mode of life appears less likely considering the great horizontal extension of the burrows.Detailed discussion of this ichnogenus sees Osgood(1970),Pemberton and Frey(1982)and Keighley and Pickerill(1995).
Planolites beverleyensis Billings,1862(Fig.9A)
Long,straight to widely curved,horizontal,cylindrical simple burrows,preserved as full relief.The fill is identical to the surrounding matrix.A lining is absent.The diameters of the burrows vary between 0.1 cm,0.2 cm,0.3 cm,and 0.5 cm.The longest extension observed is 26 cm.
Planolites is one of most common ichnotaxa living in low-energy,quiet habitats and also occurring in the aftermaths of major Phanerozoic mass extinctions(e.g.,Chen et al.,2011,2015).Most likely,these burrows have been produced by deposit-feeding organisms(e.g.,Pemberton and Frey,1982).
Taenidium cf.diesingi(Unger,1850)(Fig.9B)
Taenidium cf.diesingi is a slightly winding horizontal full relief ribbon that lacks a lining and differs from Taenidium serpentinum chiefly by its fill that is less arcuate than in the latter and is composed of regular alternations of pads that are 0.1—0.2 cm thick.The burrow diameter is 0.9—1.0 cm;in a single case it was 0.5 cm.As in T.serpentinum,the two types of sediment fill differ in their clay content.According to Uchman(1999),Taenidium satanassi D'Alessandro and Bromley,1987 is a junior synonym of T.diesingi.As the preservation quality of the trace fossil precluded the observation of a characteristic feature of T.diesingi,the presence of pellets,the specimens are placed in the ichnospecies with reservation.Ethological and trophic category:as in T.serpentinum.
Taenidium serpentinum Heer,1877(Fig.9C—E)
Taenidium serpentinum occurs as epichnial,horizontally winding cylinders,0.9—1.2 cm in diameter(average:1 cm),without lining,preserved in positive full relief in six out of 15 beds investigated.Burrows commonly cross each other.The arcuate menisci,which consist of the same material as the surrounding matrix,are 0.4—0.6 cm thick.The individual pads are separated by sub-millimetric to mm-thick softer arcuate laminae that weather more easily,probably due to their higher clay content.
T.serpentinum is interpreted as the product of a deposit-feeding worm-like organism(e.g.,D'Alessandro and Bromley,1987;Keighley and Pickerill,1994)and thus belongs to the fodinichnia.
Teichichnus cf.patens Schlirf,2000(Fig.9F)

Fig.9 Trace fossils from the upper surfaces of amalgamated grainstone beds. A — Planolites beverleyensis Billings, 1862; B — Taenidium cf.diesingi (Unger, 1850); C-E — Taenidium serpentinum Heer, 1877; F — Teichichnus cf. patens Schlirf, 2000.
Teichichnus patens differs from the much more common Teichichnus rectus by common branching.The specimen in Fig.9F clearly shows retrusive spreiten and angular changes in the long axis of the burrow.As well-developed branches are not seen the identification istentative.The some what zigzag course of the burrow also resembles that of Treptichnus.One specimen,which is 2 cm in diameter,could be followed for 40 cm.It is preserved as full relief.The final tube is oval in cross-section due to compaction.The height of the spreite appears to be at least that of the burrow diameter.
Teichichnus cf.patens has been produced by a deposit-feeding organism and can be classified as fodinichnion(Schlirf,2000).
?Teichichnus rectus Seilacher,1955(Fig.10A)
The simple,straight,cylindrical burrows,preserved in full relief,exhibit a slight downward directed curvature in a vertical plane.The burrow diameter is 1.2 cm,and the longest extension observed is 23 cm.The retrusive spreiten is fairly short,and it is for this reason that the identification as T.rectus is questionable as several other ichnotaxa may contain a similar spreiten-like feature(e.g.,Thalassinoides;Fürsich,1974: fig.23).
?T.rectus is a fodinichnion produced by an organism that systematically mined the sediment for food thereby creating a spreite.
Thalassinoides suevicus(Rieth,1932)(Fig.10B)
Thalassinoides suevicus occurs as horizontal cylindrical burrows,which are characterized by Y-shaped branching and may form networks that occur in two size classes:Type A consists of burrows 1.5—2 cm in diameter,while type B the burrow diameter ranges generally from 3 cm to 4.5 cm,on one bedding plane from 5 cm to 6 cm.Some of the negative epireliefs are enlarged to a diameter of 10 cm by erosion.In both types small parts of the wall may be indistinctly pelleted.The burrow fill is usually intraclastic and bioclastic marl.
There is general agreement that the burrows are produced by crustaceans and served as domichnia.In analogy of their modern counterparts the producers most likely were scavengers/omnivores(e.g.,Bromley,1996;Frey et al.,1984;Giannetti et al.,2007).
?Thalassinoides isp.A(Figs.10C,D,and 11)
The small?Thalassinoides isp.A is characterized by an irregularly curved course of the cylindrical burrows and numerous branchings resulting in horizontal networks.The burrows are 1 cm in diameter,occasionally exhibit meniscate back fill structures,and are usually filled with the same material as the surroundingmatrix.Commonly,a thin liningis developed.The trace fossil occurs as full relief or as negative epirelief.

Fig.10 Trace fossils from the upper surfaces of amalgamated grainstone beds.A—?Teichichnus rectus Seilacher,1955;B—Thalassinoides suevicus Rieth,1932.Hammer length of 33 cm for scale;C-D—?Thalassinoides isp.A.
Some of the branching,especially where only short side branches are developed(Fig.11),may be of the primary successive style(Bromley,1996: fig.8.8c).This is supported by the occasionally observed meniscate back fill.If true,this would not fit the diagnosis of Thalassinoides,which is an open burrow system.For this reason,the trace fossil is referred to Thalassinoides only with reservation.Alternatively,the ichnotaxon might be a variant of Taenidium,based on the presence of meniscate back fills and its irregularly curved course.
Sinusichnus Gibert, 1996 displays branching preferably occurring on the convex side of sinuous meanders and does not display meniscate filling(Knaust et al.,2016).Its type ichnospecies,Sinusichnus sinuosus Gibert,1996,is a horizontal burrow system with sinusoidalelements and multiple branching.It is somewhat similar,but our material lacks the retrusive spreiten mentioned by Belaústegui et al.(2015),and the sinuosity is far less developed than in S.sinuosus.
The burrows have been described as Thalassinoides suevicus type C by Fürsich(1998).They are also quite common in event beds in the basal-most part of the overlying Chari Formation.As in the case of T.suevicus,the burrows are domichnia and the producers were most likely scavenging/omnivorous crustaceans.
5. Data analysis
5.1. Preservation and abundance
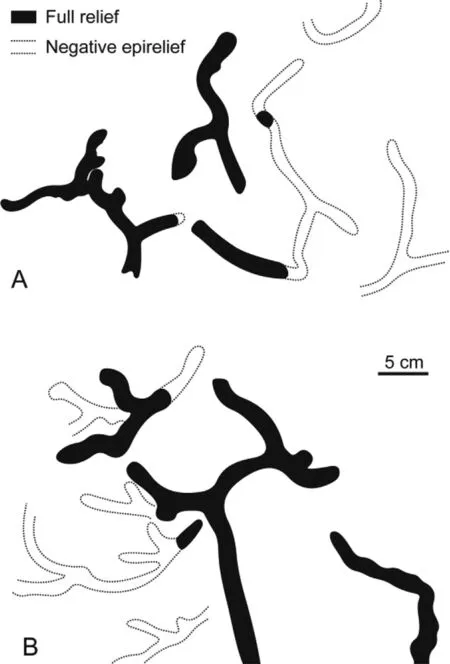
Fig.11 A—B — Sketches of?Thalassinoides isp.A showing the curved nature of the tunnels and the multiple branching.Some branching appears to be of the primary successive style.Bedding plane view.
Within the investigated sedimentary succession(Fig.5),trace fossils occur mainly on the top of individual grainstone beds and in the intraclastic—bioclastic silty marl interbeds.In the latter,bioturbation is so intense that no individual traces can be discerned.Compaction and the unconsolidated nature of the interbeds in addition made it impossible to identify any ichnotaxa.In the grainstones,in contrast,trace fossils are well preserved but sparsely distributed,except for some few patches,in which the burrow density is distinctly higher.The trace fossils are preserved as negative epireliefs or as full reliefs.No lower bed surfaces were available for investigation.Vertical components(e.g.,shafts)are rarely seen in bed cross-sections;instead,most traces are horizontally aligned,although vertical elements must have been present in several of the ichnotaxa such as Ophiomorpha,Thalassinoides,Gyrophyllites and Dactyloidites.Apparently,the vertical elements were far less common and/or conspicuous than horizontal ones and often might have been removed by erosion.
Considering that all traces were produced within the sediment,their preservation on the top of the grainstone beds requires an explanation.The reason is that their corresponding colonization horizon from which they extended downwards up to or into the top of a grainstone bed was situated within the overlying silty marl unit.
5.2. Ichnodiversity
Altogether 18 ichnotaxa have been recorded from the section(Table 1).Most bed surfaces show a moderate diversity(4—7 ichnotaxa).The highest diversity with ten ichnotaxa is found on the surface of bed 10,the lowest diversity(0)occurs in bed 14,followed by bed 15(1 ichnotaxon).In the highly bioturbated marl interbeds the ichnofabric probably has been produced by a number of different organisms,but as discussed above,its ichnodiversity cannot be reconstructed.
5.3. Behavioural and trophic composition
Most of the ichnotaxa(61%;Table 1)can be classified as fodinichnia(feeding structures produced more or less in the same place)or pascichnia(combined feeding and locomotion structures).Thirtythree percents are predominantly domichnia(dwelling burrows),partly in combination with a deposit feeding behaviour and thus are secondarily also fodinichnia.Cubichnia(?Bergaueria;6%)are rare.
Three-fourth(78%)of the ichnotaxa were produced by organisms with a deposit-feeding mode of life.They probably include both,selective and non-selective feeding habits;two ichnotaxa(Ophiomorpha nodosa,Bergaueria;11%)were produced bysuspensionfeeders,whilst the producer of Thalassinoides suevicus probably was a scavenger or omnivore.
5.4. Ichnoassemblages
Ophiomorpha nodosa—Thalassinoides suevicus ichnoassemblage—The distribution of ichnotaxa and their diversity and abundance within the ten grainstone beds containing trace fossils is not uniform.In bed 14,trace fossils are absent and in beds 1,3,and 15 they are represented only by one to four ichnotaxa.The latter define a low-diversity ichnoassemblage being dominated by large T.suevicus and/or O.nodosa.The assemblage consists mainly of horizontal branching tunnel systems inhabited by suspensionfeeders(O.nodosa)and scavengers/omnivores(T.suevicus,?T.isp.A),and served as domichnia/fodinichnia.Vertical shafts,which are an essential part of the burrow systems,are rare.This suggests that only the deepest parts of the three-dimensional system are preserved. Pascichnia are represented by rare Taenidium.
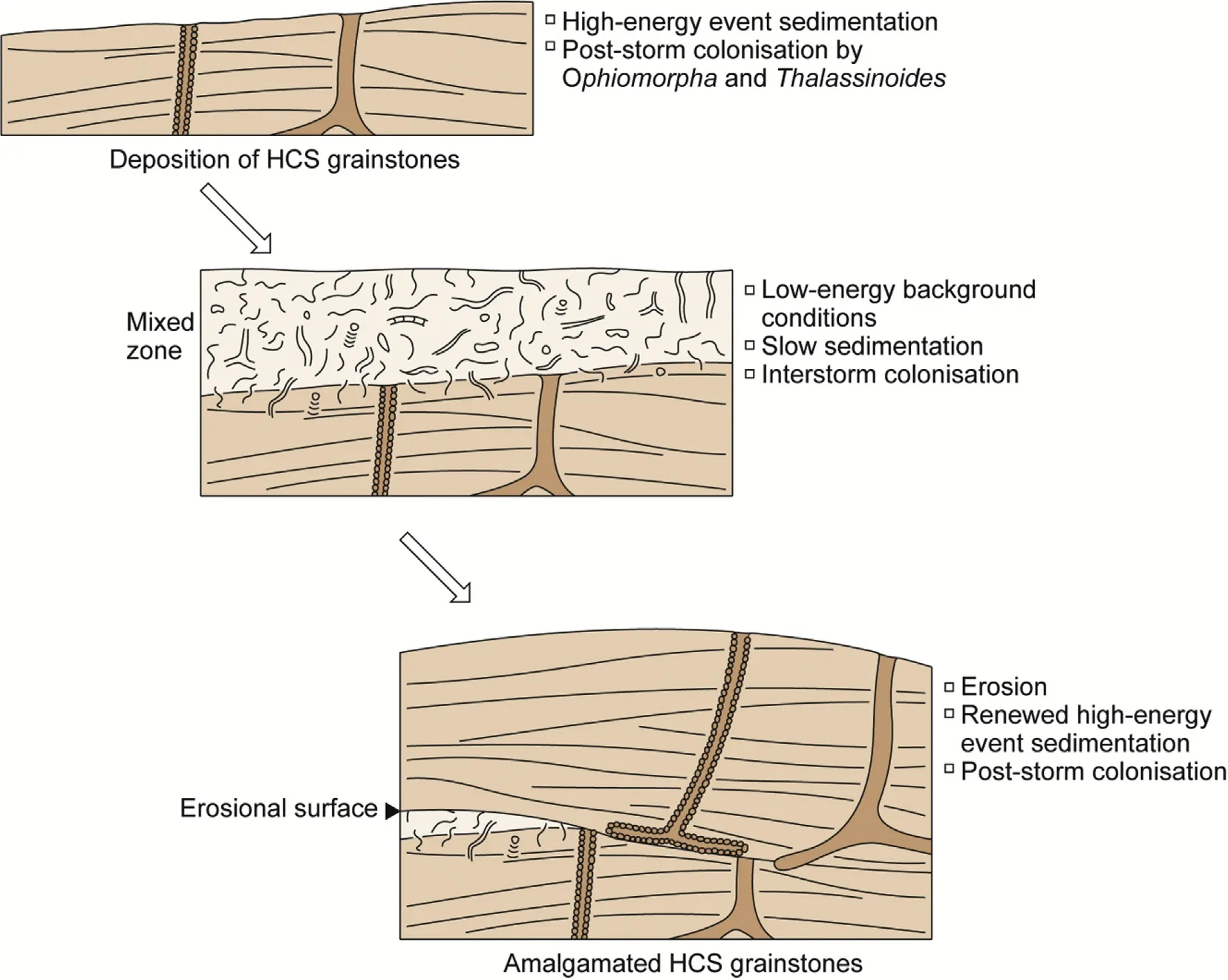
Fig.12 Model explaining the preservation of traces of deposit-feeders at the top of HCS grainstones formed during high-energy events.The co-occurrence of traces indicative of high-energy and low-energy conditions is due to time averaging.
?Thalassinoides sp.A—Taenidium—Bolonia lata ichnoassemblage—This assemblage occurs in beds 7,9,and 10 and is characterized by a comparatively high diversity(10 ichnotaxa in bed 10)and a wider range of behavioural classes than the preceding assemblage:domichnia/fodinichnia (T.suevicus,?T.isp.A), pascichnia(Taenidium,B.lata)and fodinichnia(e.g.,Gyrophyllites, Dactyloidites)are present.The prevailing trophic mode of the producers was depositfeeding.The ichnotaxa are concentrated on bedding planes and occasionally form small clusters(e.g.,Gyrophyllites,Dactyloidites).
Ancorichnus ichnoassemblage—This assemblage is similar in composition to the preceding assemblage but lacks radial feeding burrows(Gyrophyllites,Dactyloidites).The dominant ichnotaxon is Ancorichnus,which is abundant in the troughs of current ripples locally preserved on top of bed 12(Fig.6A and B)and which can be interpreted as pascichnion of a deposit feeding organism.The diversity is reasonably high(7 ichnotaxa).
6. Discussion
Based on sedimentological evidence,the top part of the Sponge Limestone member has been deposited in an upper shoreface environment,within the reach of storm waves,but below the fair-weather wave-base(Fig.12).Storm waves were strong enough to erode much,and often all of the interstorm sediments,leaving only lenticular relicts.Sedimentation was characterized by two regimes:(1)rapid,storm-wave induced deposition of calcarenites with hummocky cross-stratification and(2)low-energy conditions with comparatively low rates of deposition of fine-grained sediments.During the latter regime ample time was available for benthic organisms to colonize the muddy sea floor,which in due time became thoroughly bioturbated.In the former regime,rapid sedimentation of the decimeter-thick units was largely responsible for the scarcity of trace fossils.In addition,the content of particulate organic matter in the well-washed sediments probably was fairly low rendering them unattractive for deposit-feeding organisms.Frequent phases of erosion led to amalgamation of beds produced by storm waves(cf.Pemberton et al.,1992).
6.1. Trace fossil diversity
In many studies,trace-fossil diversity is regarded as an important feature to evaluate environmental parameters and stress conditions.However,the diversity of trace fossils inherently differs from that of body fossils.The preservation potential of trace fossils strongly depends on the burrow density:within the totally bioturbated mixed layer,it is often impossible to recognize individual ichnotaxa,although the diversity may be high.Another very important aspect is the presence of internal surfaces.Nearly all trace fossils that we observe were made within the sediment.Their preservation potential is highest at internal surfaces.Trace fossil diversity is also related to the degree of erosion.In the present case,trace fossil diversity is lowest where erosion removed the background sediments and the top parts of the grainstone storm beds so that only deep-tiertrace fossils(Ophiomorpha,Thalassinoides)are preserved(see also Wetzel and Aigner,1986).Last not least,trace fossil diversity strongly depends on the nature of the substrate.In the unconsolidated fine-grained sediment representing background conditions trace fossil diversity was arguably much higher than in the well cemented grainstones,but no information can be retrieved from the former.A final point limiting the ecological meaning of ichnodiversity is the noncontemporaneity of trace fossils with different vertical extensions.In the present study this is illustrated by the deep-burrowing producers of Ophiomorpha and Thalassinoides that may have burrowed through a number of beds(multilayer colonisers;Uchman,1995)and occur associated with traces that were excavated during an earlier time interval and reflect different environmental conditions. Ichnodiversity is also related to the time available for colonisation of the substrate,a feature trace fossils share with body fossils.In conclusion,ichnodiversity is beset with a number of pitfalls,which greatly limit its use in simple ecological interpretation of trace fossil suites.
6.2. Ichnocoenoses
The term ichnocoenosis has been coined in analogy to the term biocoenosis.It is defined as traces produced by members of an endobenthic community(Bromley,1996;Radwaénski and Roniewicz,1970)and thus implies contemporaneity.This contemporaneity is commonly not true of assemblages of trace fossils,even if they occur on the same bedding plane.This point is also illustrated by the top of the HCS grainstones in the present example where post-storm traces occur on the same bedding plane as traces produced during the preceding interstorm phase(Fig.12).We advocate to discontinue the use of ichnocoenosis and to use more neutral terms such as ichnoassemblage.
6.3. Ichnofacies
The trace fossils of the top part of the Sponge Limestone member belong to the Cruziana ichnofacies.This ichnofacies is characteristic of shelf environments below the fair-weather wave-base where fluctuations in turbulence levels are not unusual,especially in the upper shoreface.As a result several ichnoassemblages may replace each other.In the present case the low-diversity Ophiomorpha nodosa—Thalassinoides suevicus ichnoassemblage became established during brief post-storm interludes,whereas during the extended background sedimentation moderately diverse ichnoassemblages with a number of ichnoguilds(groups of ichnospecies sharing resources in a similar way)but dominated by fodinichnia and pascichnia prevailed.
6.4. Time-averaging
Time-averaging appears to be a pervasive feature of trace fossil assemblages.Where environmental conditions remain uniform does not pose a major problem for their interpretation.If environmental conditions change,the interpretation of the distribution pattern of trace fossils at the boundary between two environmental states requires special care.In our example,the two environmental states are(a)brief,high-energy storm conditions and corresponding rapid sedimentation of sand-sized carbonate particles with low Corgcontent,and(b)distinctly longer-lasting low energy conditions with lower rates of sedimentation of Corg-rich finer-grained particles.At the end of state(a)a brief colonisation window existed for traces of basically suspension-feeding (Ophiomorpha) and scavenging/omnivorous(Thalassinoides)organisms,which constructed vertical burrows turning downwards into horizontal networks.Due to the brief duration of this phase,the trace fossil density remained low.During environmental state(b)ample time was available for bioturbation by a benthic community dominated by deposit-feeders.Their burrows extended occasionally into the top of the underlying carbonate sand(now grainstones),but due to its low food content exploitation of the sand-sized sediment remained sporadic or just affected the top surface(Fig.12).This led to the inconsistency that the high-energy stormwave-produced sediments are characterized by a trace-fossil assemblage largely reflecting low-energy conditions.Thus,we not only have the ichnotaxa of the two environmental states preserved on the same bedding plane but also ichnotaxa of state(b)preserved in rocks produced during state(a).
Such mixing of traces reflecting different environmental conditions and time-averaging(producing a socalled heterocomposite ichnofacies sensu Savrda,2016)does not need to involve major vertically extended burrows.Ekdale and Bromley(1991)discussed the superposition of different ichnotaxa reflecting different substrate conditions in the Maastrichtian chalk of Denmark.Another classical example of the superposition of trace fossils that reflect different substrate conditions are hard grounds,in which the increasing compaction and hardening is documented by the change from a soft ground trace fossil assemblage to a firm ground and finally hard ground assemblage(e.g.,Fürsich,1978).Thus,wherever environmental conditions fluctuate, either regularly or episodically,there is a high likelihood that the corresponding trace-fossil assemblages are highly time-averaged.As a result they commonly occur in rocks with contrasting sedimentological signatures.This should be taken into account when high-resolution environmental analyses of trace fossils are attempted.
7. Conclusions
Extensive bedding plane exposures of grainstones at the top of the Sponge Limestone member,Patcham Formation,of the Jhura Dome in Kachchh,Gujarat,India,contain low-density,but moderately diverse trace-fossil assemblages of the Cruziana ichnofacies.The hummocky cross-stratified grainstone beds are amalgamated or are separated by thin,lenticular intraclastic—bioclastic silty marl interbeds,which are so highly bioturbated that individual ichnotaxa can no longer be recognized.In most grainstone beds the majority of the ichnotaxa have been produced by deposit-feeding organisms,whereas ichnotaxa made by suspension-feeders play only a subordinate role.As the trophic composition of benthic faunas is clearly related to the level of water energy,the dominance of deposit-feeders reflects low-energy conditions,which is in contrast with the high-energy character of the grainstone beds.The sedimentary succession can be interpreted as having been deposited under a proximal storm-wave regime characteristic of the upper shoreface.Bioturbation took place during a brief,comparatively high-energy post-storm phase,when traces such as Ophiomorpha and Thalassinoides with a large vertical extension were produced. During the following interstorm phase,low-energy conditions prevailed and the background sediment became thoroughly bioturbated by deposit-feeders,with some of the burrows extending into the top part of the underlying grainstones.Erosion during the next storm event frequently removed the background sediments.The incongruous association of sediments reflecting high water-energy and trace fossils indicative of comparatively quiet-water conditions is a case of time-averaging that shows that trace fossils may document environmental conditions drastically differing from those of the rocks in which they occur.
Acknowledgements
We acknowledge Valsamma Fürsich for assistance in the field.A visit of Alfred Uchman to the field area was supported by the Jagiellonian University(DS funds).Dhirendra K.Pandey acknowledges financial assistance for the fieldwork by DST(project EMR/2015/001574)and Matthias Alberti financial support by the Alexander von Humboldt Foundation.We appreciate the critical remarks by Profs.Yi-Ming Gong,Bin Hu,and Kantimati Kulkarni.
 Journal of Palaeogeography2018年1期
Journal of Palaeogeography2018年1期
- Journal of Palaeogeography的其它文章
- Late Triassic sauropodomorph and Middle Jurassic theropod tracks from the Xichang Basin,Sichuan Province,southwestern China:First report of the ichnogenus Carmelopodus
- Oxygenation in carbonate microbialites and associated facies after the end-Permian mass extinction:Problems and potential solutions
- Evolution of sedimentary facies and fossil communities in the Middle Permian Maokou Formation in Zigui County,Hubei Province,South China
- A redescription of the ichnospecies Koreanaornis anhuiensis(Aves)from the Lower Cretaceous Qiuzhuang Formation at Mingguang city,Anhui Province,China
- Geologic-seismic models,prediction of shallow-water lacustrine delta sandbody and hydrocarbon potential in the Late Miocene,Huanghekou Sag,Bohai Bay Basin,northern China
- Deeply concealed half-graben at the SW margin of the East European Craton(SE Poland)—Evidence for Neoproterozoic rifting prior to the break-up of Rodinia