
榛属ycf1基因生物信息学分析
2018-08-17 03:16卫永乐温志芳张杰伟黄武刚兰彦平程丽莉曹庆昌胡广隆
山西农业科学 2018年8期
卫永乐,温志芳,刘 芳,张杰伟,黄武刚,兰彦平,程丽莉,曹庆昌,胡广隆
(1.运城农业职业技术学院,山西 运城 044000;2.北京市农林科学院,北京 100093)
叶绿体在植物细胞中具有与光合作用密切相关的一整套基因。鉴于叶绿体基因组的大小、结构和基因型相对保守性,其已成为植物系统发育研究和识别物种的有力工具[1]。目前,某些叶绿体基因的功能已被研究,但有相当一部分基因是未知的,例如一大类基因,即开放阅读框(ORF)。它的DNA序列所编码的特定蛋白质名称和功能仍在摸索中。由于ORF命名方法有诸多不妥,因此,学术界提出了一种新的YCF(假设叶绿体开放阅读框)命名方法。例如,小麦的ORF38与菠菜ORF33的氨基酸序列相似性很高,因此都命名为ycf8基因[2]。
叶绿体DNA(cpDNA)序列已被分类学家广泛用于重建植物系统发育[3],然而,相对较少的叶绿体基因组区域被用于系统发育研究[4-5]。蛋白质编码基因容易比对,但通常更加保守,缺乏足够的变异来解决种间和种内关系[6-7]。比较基因组的研究表明,叶绿体体基因 ycf1,(hypothetical chloroplast open reading frame 1)可以比matK基因变异度更高[8]。ycf1是叶绿体基因组中第二长的基因(仅次于ycf2),并存在于迄今测序的几乎所有植物叶绿体基因组中[9]。YCF1蛋白的功能是未知的。尽管如此,DRESCHER等[10]研究已经证明,YCF1对植物生存至关重要。ycf1阅读框通常跨越倒置重复区(IR)和小单拷贝(SSC)区域的边界,这在叶绿体基因中是比较特殊的[9]。ycf1基因在系统发育研究中的效用最近才开始探索。NEUBIG等[11]研究表明,ycf1因为其保守的阅读框,是相对容易扩增和比对的。此外,在兰花分类学研究中,ycf1具有类似或仅低于细胞核核糖体DNA内转录间隔区(ITS)的高度变异性,比matK有更优异的分辨率。
榛属植物约有20种,分布在北美洲、欧洲和亚洲;其中,我国有7种及3个变种,主要在东北、华北、西北和西南地区[12]。该属植物的种子含油量大,在东北、华北地区当作油料;坚果可食用,为北方地区常见干果。乔木树种木质硬,用于家具制作及建筑业。二代测序技术的应用加快了榛属叶绿体全基因组研究,榛属叶绿体全基因组的发表[13]为榛属植物叶绿体基因奠定了生物信息学分析与鉴定的基础。
本研究充分利用榛子叶绿体基因组数据库,以榛属ycf1基因家族成员为研究对象,着重分析其基因结构、跨膜区的结构特征和编码区的保守结构域,并与拟南芥及相邻的铁木属经过多重序列比对进行进化和分类分析,旨在为克隆榛子ycf1基因及进一步探索YCF1蛋白的功能奠定坚实的基础。
1 材料和方法
1.1 榛属ycf1基因组、cDNA和蛋白序列的来源
榛属、铁木属及拟南芥的ycf1基因及蛋白序列分别下载自 NCBI网站(https://www.ncbi.nlm.nih.gov)和 TAIR 数据库(http://www.arabidopsis.org)。利用NCBI的Blast P检索及CDS软件(http://www.ncbi.nlm.nih.gov/cdd)预测所选蛋白有无YCF1结构域,判断其是否属于该基因家族。
1.2 榛属ycf1基因家族结构分析
利用数据库,获取榛属中ycf1基因家族成员的基因注释。
1.3 榛属ycf1基因家族二级结构及跨膜区结构的预测
ycf1基因家族各成员所编码蛋白质氨基酸的相似性分析[14]、α-螺旋、β-折叠及无规则卷曲等二级结构分析[15]、跨膜区结构[16]分析等分别采用Clustal Omega(http://www.ebi.ac.uk/Tools/msa/clustalo/),SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)和 TMHMMServer v.2.0(http://www.cbs.dtu.dk/services/TMHMM-2.0/)等软件。
1.4 榛属ycf1基因家族的保守结构域分析
利用软件 MEME(http://meme-suite.org/)分析拟南芥、铁木属和榛属ycf1基因蛋白的保守结构域[17]。
1.5 榛属ycf1基因蛋白三级结构的预测
榛属和铁木属YCF1蛋白质空间结构模型的同源建模[18]由三级结构预测软件 Swiss-Model(https://swissmodel.expasy.org/)的自动建模功能完成。
1.6 榛属ycf1基因蛋白系统进化树的构建
拟南芥、铁木属和榛属的ycf1基因蛋白序列的比对利用Clustal X(2.0)[19]软件进行;将比对结果用MEGA 6.0[20]软件采用Neighborjoining法(bootstrap值设定为1 000)产生系统进化树。
2 结果与分析
2.1 榛属ycf1基因家族成员的评价
利用NCBI数据库提供的Blast程序进行Blast P检索(图1),得到3个榛属ycf1基因和1个铁木属ycf1基因。用保守域预测软件CDS验证了ycf1基因保守结构域的存在,初步确定榛属中3个ycf1基因分别来自华榛、平榛、欧榛;另外,在同科相邻的铁木属的天目铁木中也确定了1个ycf1基因作为对比研究基因,即:YP009331532.1(华榛),AOZ20414.2(欧榛),YP009318292.1(平榛),YP009179114.1(天目铁木)。这几个基因均没有内含子。
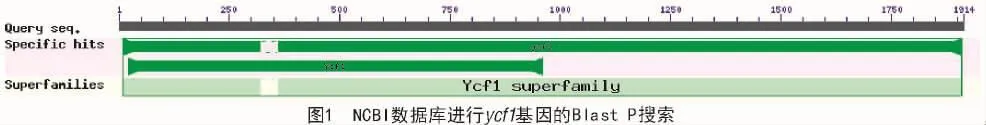
2.2 榛属ycf1基因家族的结构分析
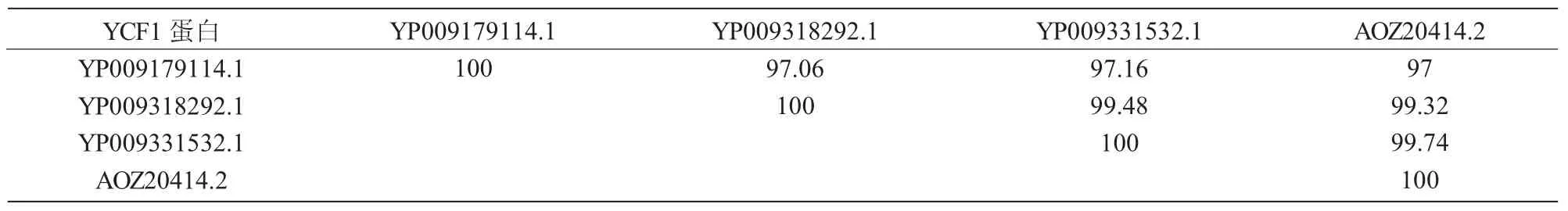
表1 YCF1蛋白氨基酸相似性分析%
ycf1基因家族各成员编码蛋白质氨基酸的相似性分析结果表明,欧榛YCF1蛋白和华榛YCF1蛋白之间的相似性最高,达99.74%;欧榛YCF1蛋白和天目铁木YCF1蛋白之间的相似性最低,为97.00%(表1)。二级结构预测分析结果表明,榛属YCF1蛋白中 α-螺旋的比例最高(34.90%~35.89%),其次为无规则卷曲(29.91%~30.25%),再次为β-折叠(25.29%~26.15%),β-转角最低(8.73%~9.04%)(表2)。榛属YCF1蛋白的跨膜区结构分析结果表明,榛属及铁木属YCF1蛋白均含有跨膜区(图2)。

表2 榛属及天目铁木YCF1蛋白的二级结构信息 %
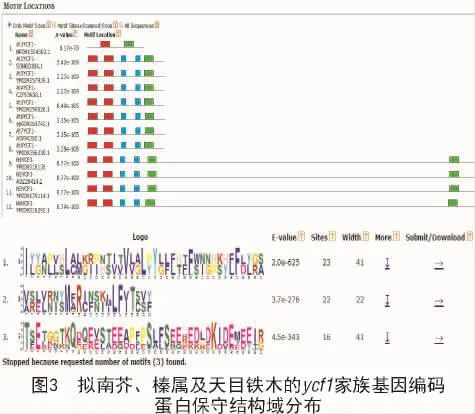
利用保守结构域预测软件MEME分析了拟南芥、铁木属和榛属ycf1基因家族的保守结构域,结果显示,拟南芥、榛属和铁木属ycf1基因家族包含3个保守结构域。其中,基序1的相似度为2.0e-625;基序2和基序3的相似度分别为3.7e-276和4.5e-343(图3)。除了拟南芥ycf1基因At1YCF1,无论在桦木科的榛属和铁木属中,还是在十字花科的拟南芥中,ycf1基因家族均含有3个结构域(图3)。
本研究通过Swiss-Model对榛属及铁木属YCF1蛋白进行同源建模,只发现有5%的序列能与已知模板匹配,即只与转运蛋白AcrB的一个亚基同源。从图4可以看出,Swiss-Model预测的榛属及铁木属YCF1蛋白亚基三级结构的空间结构非常相似,暗示不同桦木科YCF1蛋白在生物学功能上存在相似性。

2.3 榛属YCF1家族蛋白质相似性和系统进化树分析
利用软件MEGA 6.0对8个拟南芥YCF1蛋白、4个榛属及铁木属YCF1蛋白的序列进行系统进化树分析,结果表明,这12个YCF1蛋白序列基本按照物种聚类。YCF1蛋白系统进化树大致上按照科别进行聚类,推测YCF1蛋白在桦木科和十字花科中生物学功能不尽相同。华榛YCF1(YP009331532.1)与欧榛 YCF1(AOZ20414.2)聚类在一起,暗示华榛YCF1可能具有与欧榛YCF1相似的进化过程和生物学功能(图4)。YCF1蛋白在桦木科的进化并没有完全按照属聚类在一起,相对于平榛,天目铁木的YCF1蛋白与欧榛及华榛的关系更近(图 5)。

3 讨论
YCF1具体编码的蛋白质名称和功能仍在探索中。作为叶绿体基因组中2个最大的开放阅读框架之一,ycf1常被认为是失去功能的假基因[21-23]。然而,YCF1的敲除试验显示,YCF1是细胞存活所必需的[24]。杨小飞等[25]研究发现,核基因组编码的RNA结合蛋白PBR1,通过调控拟南芥YCF1在叶绿体中的翻译效率,进而调节3个光合作用复合体的协同生成及稳定性的维持。目前,榛属ycf1基因家族具体功能尚未鉴定。本研究通过生物信息学鉴定榛属ycf1基因家族3个基因均含有跨膜区,均含有3个结构域,表明榛属中不同种间ycf1基因家族中重要结构域存在高度保守性。另外,除了拟南芥ycf1基因编码蛋白At1YCF1,无论在桦木科的榛属和铁木属中,还是在十字花科的拟南芥中,ycf1基因家族均含有3个结构域。推测YCF1的3个结构域在植物诸多生理过程中发挥相似的作用。
进化分析结果表明,华榛YCF1(YP009331532.1)与欧榛YCF1(AOZ20414.2)聚类在一起,暗示他们可能具有相似的进化过程和生物学功能;YCF1蛋白在桦木科的进化并没有完全按照属聚类在一起,相对于平榛,天目铁木的YCF1蛋白与欧榛及华榛的关系更近。基因的进化并非完全与种属分类进化一致,同样的情况出现在兰花磷酸肌醇特异性磷脂酶C基因家族的进化中[26]。由此可见,榛属YCF1各成员的生物学功能还需要进一步分析研究。
4 结论
榛属植物叶绿体基因组中ycf1家族基因,均不含有内含子。榛属YCF1蛋白均含跨膜区和3个高度保守结构域。进化分析结果表明,华榛ycf1与欧榛ycf1可能具有相似的进化过程和生物学功能。
猜你喜欢
国际放射医学核医学杂志(2020年4期)2020-07-27
生物信息学(2020年1期)2020-05-16
少儿科技(2019年9期)2019-09-10
广州大学学报(自然科学版)(2019年1期)2019-05-07
上海农业学报(2017年3期)2017-04-10
广西林业科学(2016年1期)2016-03-20
天津科技大学学报(2016年1期)2016-02-28
现代检验医学杂志(2015年2期)2015-02-06
天然产物研究与开发(2014年6期)2014-04-27
植物营养与肥料学报(2014年1期)2014-03-11
