
结核分枝杆菌休眠存活调节子蛋白Rv2029c促进大肠埃希菌生长
2018-08-11 01:19:02王永祥张本忠李豪杰何文华高小玲
检验医学与临床 2018年15期
王永祥,张本忠,吴 聪,李豪杰,何文华,高小玲
(1.兰州大学公共卫生学院,兰州 730000;2.甘肃省人民医院转化医学研究所,兰州 730000)
据WHO估计,全球约有1/3的人口是结核分枝杆菌潜伏感染患者,其中有5%~10%的人会发展成活动性肺结核[1-2]。潜伏期结核分枝杆菌存在于肉芽肿组织中,这是一种低氧、高一氧化氮、营养物质匮乏的微环境[3]。结核分枝杆菌在此环境中处于休眠期,生长速度非常缓慢,也正是因为如此,结核分枝杆菌逃避了机体免疫系统的作用,从而存活下来[4-5]。进一步的研究发现,结核分枝杆菌基因组存在着一组包含48个基因的休眠存活调节子(DosR),这48个基因编码的蛋白控制着结核潜伏期的生理过程[6]。肉芽肿组织中的结核分枝杆菌存在着依赖氧压的细胞和生化动力学机制,能够使其快速适应微环境改变的能力,在此过程中结核分枝杆菌DsoR基因起着至关重要的作用[7-8]。与活动期结核分枝杆菌相比,DosR区域编码的抗原能够引起潜伏期结核患者产生更强烈的免疫反应[9],这表明DosR基因很可能在潜伏期特异性表达。有研究报道,DosR基因编码的许多蛋白有助于结核分枝杆菌利用其他碳源途径获得能量,如乙醛酸代谢、硝基还原和脂肪酸代谢等[10-11]。探索DosR基因生物学功能有助于进一步认识结核分枝杆菌潜伏期的生理调控过程,从而有针对性地进行检测,实现早发现、早诊断、早治疗的目的,对于预防活动性结核的发生至关重要。
1 材料与方法
1.1仪器与试剂 本实验试剂结核分枝杆菌H37Rv株,载体pET30a由本实验室保存。大肠埃希菌BL21(DE3)感受态细胞购自南京诺唯赞公司;质粒提取试剂盒购自北京天恩泽公司;胶回收试剂盒购自Zymo公司;各种限制性内切酶购自Thermo公司;DNA聚合酶和连接酶购自TaKaRa公司;异丙基硫代-β-D-半乳糖苷(IPTG)购自Sigma公司。美国Biorad酶标仪。
1.2方法
1.2.1结核分枝杆菌DNA的提取 结核分枝杆菌DNA的提取依据已报道的文献进行[12]。具体方法如下: 挑取结核分枝杆菌菌落,置于装有200 μL生理盐水的1.5 mL离心管中,80 ℃煮沸灭活30 min,12 000 r/min离心15 min后,加入蛋白酶K和溶菌酶充分裂解结核分枝杆菌,然后用酚-氯仿抽提法获取结核分枝杆菌基因组,无水乙醇沉淀双链DNA,干燥后TE缓冲液(50 μL)溶解DNA。
1.2.2引物设计 在DBGET数据库中查找结核基因Rv2029c的全基因序列,然后采用Primer5.0和Oligo6.0设计Rv2029c特异性引物序列,上下游分别插入EcoRⅠ和HindⅢ酶切位点。设计的引物序列为上游引物Rv2029F:CGG AAT CCA TGA CGG AGC CAG CGG CGT,其中CG是保护性碱基,GAA TCC是EcoRⅠ酶切位点;下游引物Rv2029R:CCC AAG CTT TCA TGG CGA GGC TTC CGG GTT,其中CCC是保护性碱基,AAGCTT是HindⅢ酶切位点。
1.2.3纯化目的基因 采用聚合酶链式反应(PCR)的方法对目的基因进行扩增,扩增条件:98 ℃变性10 s,55 ℃退火5 s,72 ℃延伸80 s,共扩增35个循环。扩增后的目的基因利用胶回收试剂盒(美国Zymo公司胶回收试剂盒,D4007)进行纯化和浓缩。具体方法如下:首先在琼脂糖凝胶上准确切取目的基因片段,然后加入凝胶溶解液,37~55 ℃孵育融化5~10 min后,过柱离心吸附,加Washing buffer洗涤2次后,用10~20 μLTE缓冲液洗脱即可。
1.2.4重组质粒的构建 重组质粒PE730a-Rv2029c的构建依据文献[12],主要概述如下:用限制性内切酶EcoRⅠ和HindⅢ对目的基因和载体pET30a进行双酶切,然后经1.2%的琼脂糖凝胶电泳后切胶回收。目的基因和载体按3∶1的比例,在DNA连接酶的作用下,16 ℃过夜连接。连接后的目的基因,导入到大肠埃希菌感受态(BL21)细胞中,4 ℃水浴30 min后,42 ℃准确热激45 s后,加入无抗菌药物的LB培养液后,细菌复苏45 min,然后将细菌均匀涂到含卡那霉素(50 μg/mL)的LB固体培养皿中,37 ℃过夜后挑取阳性单克隆菌落并提取质粒(北京天恩泽公司质粒提取试剂盒,60205),双酶切(EcoRⅠ和HindⅢ)鉴定后送南京金斯瑞公司进行测序分析。
1.2.5重组质粒pET30a-Rv2029c促生长作用研究 大肠埃希菌BL21是一种常用的基因工程表达菌株,其生长分为迟缓期、对数期、平台期和衰亡期。迟缓期大肠埃希菌适应新的环境,因此生长速度很慢,对数期营养物质充分,细菌呈对数增长,随着培养基中营养物质的不断消耗,大肠埃希菌代谢产物不断积累,培养基pH逐渐下降,大肠埃希菌生长速度变慢,细菌总数趋于稳定。大肠埃希菌的平台期与结核分枝杆菌的肉芽肿微环境相似,对细菌的生长都是一种不利的环境,以此模拟肉芽肿微环境,将结核DosR蛋白导入大肠埃希菌BL21中,观察对其生长有无影响。
取出保存的pET30a大肠埃希菌(空质粒组)和pET30a-Rv2029c大肠埃希菌(重组质粒组),在含50 μg/mL卡那霉素的5 mL LB培养液中37 ℃振荡培养过夜,记录吸光度(OD)600值。然后将菌液调整至相同的OD值并吸取600 μL菌液加至300 mL LB培养基中(含 50 μg/mL卡那霉素),37 ℃ 300 r/min振荡培养,每隔1 h取600 μL菌液,检测OD值(每次测3组OD值,每组测200 μL)。在第5个小时当2组大肠埃希菌OD值为0.45时,加入诱导剂异丙基硫代半乳糖苷(IPTG)(500 mmol/L),之后继续每隔1 h取600 μL菌液测OD值,如此连续重复测量15 h。在相同条件(相同菌种,相同生长和测量条件)下重复该实验3次。
1.3统计学处理 采用SPSS22.0对1.6实验结果进行定量数据重复测量的方差分析及t检验,比较空质粒组与重组质粒组对大肠埃希菌OD值是否产生影响,并作出生长曲线图。以P<0.05为差异有统计学意义。
2 结 果
2.1Rv2029c基因扩增结果 以提取的结核基因组为模板,在设计好的引物作用下,行PCR扩增。PCR成功扩出Rv2029c DNA,大小1 020 bp。见图1。
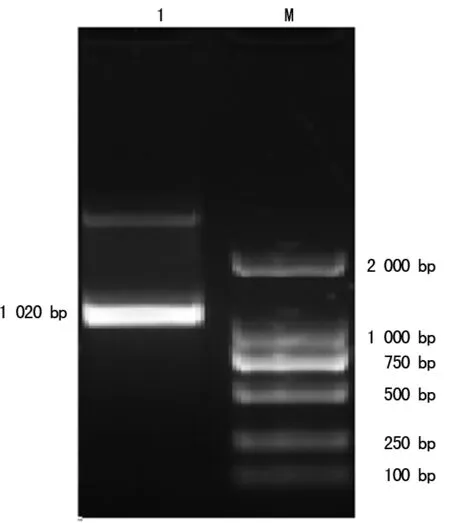
注:1为基因Rv2029c;M为DNA2000标记物
图1结核DosR基因Rv2029c PCR结果
2.2pET30a-Rv2029c重组质粒的构建与鉴定 将挑取的单克隆菌用EcoRⅠ和HindⅢ进行双酶切,酶切鉴定发现,阳性重组质粒能切出2条基因片段,分别为质粒pET30a(5 422 bp)和插入的目的基因Rv2029c(1 020 bp)。与连接片段大小吻合,可初步认为pET30a-Rv2029c构建成功。见图2。
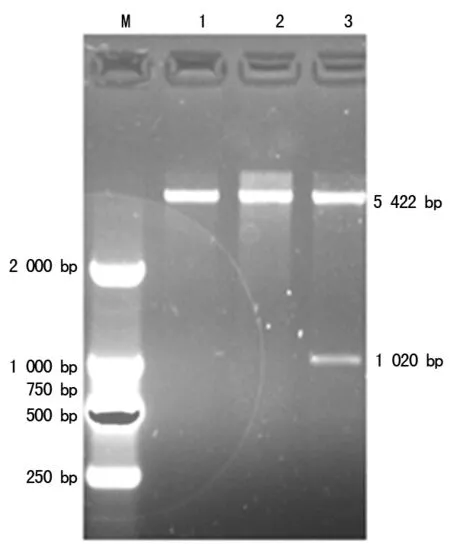
注:M为DNA标记物;1~2为阴性质粒;3为阳性质粒pET30a-Rv2029c
图2 pET30a-Rv2029c重组质粒酶切鉴定结果
2.3基因测序结果和GenBank序列比对 将酶切鉴定成功的重组质粒送往南京金斯瑞公司进行测序,测序结果与GenBank所给序列进行Blast。基因Rv2029c与GenBank中所给序列完全吻合,重复率100%,基因无突变。
2.4pET30a-Rv2029c促大肠埃希菌生长的统计学分析 将3次测量获得的资料进行重复测量的方差分析,结果显示sum值观察时间效应:不同时间OD值差异有统计学意义(F=8 351.248,P<0.05),空质粒组(pET30a)和重组质粒组(pET30a-Rv2029c)不同时间OD值差异均有统计学意义(F=3 593.681、4 125.791,P<0.05)。sum值观察处理因素效应,重组质粒组(pET30a-Rv2029c)OD值高于空质粒组(pET30a)(t=4 416.54,P<0.05)。从各时间点来看,0~5 h空质粒组和重组质粒组(pET30a-Rv2029c)OD值差异无统计学意义(t=1.372、1.220、0.894、0.935、0.787、0.395,P>0.05),6~15 h由于加入IPTG诱导了蛋白Rv2029c的表达,OD值差异有统计学意义(P<0.05);生长时间和处理因素存在交互效应(F=767.049,P<0.05)。见表1。

表1 0~15 h导入Rv2029c对大肠埃希菌OD值的影响
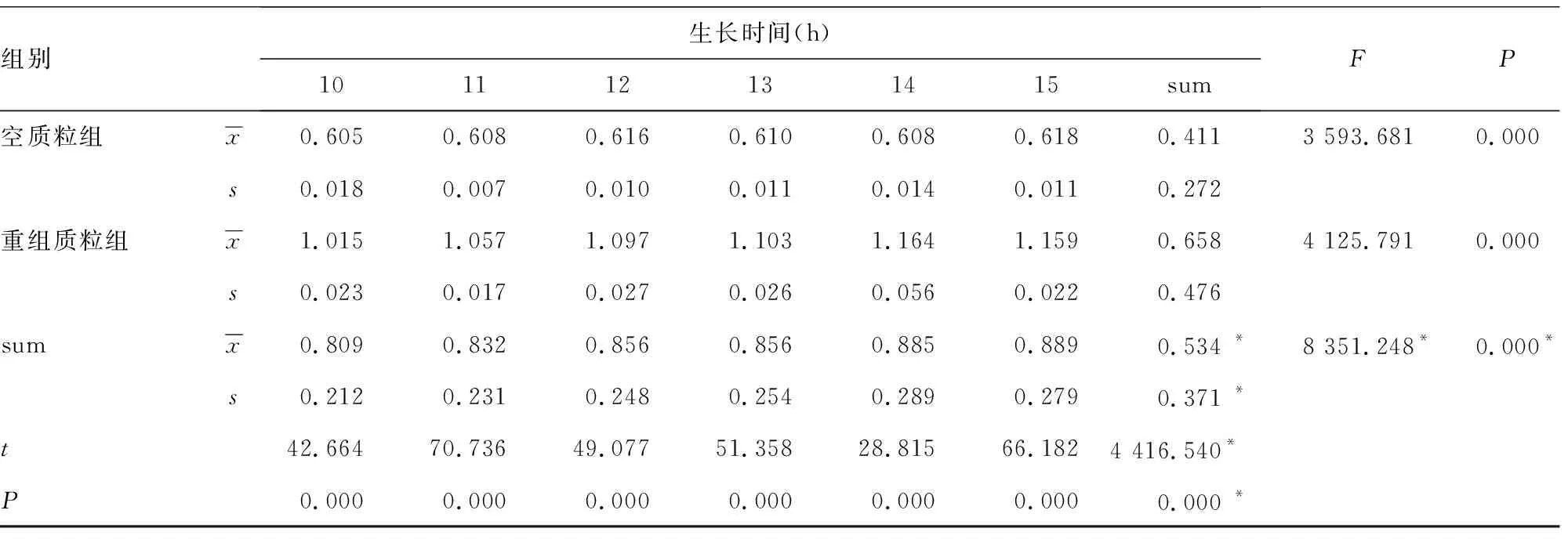
组别生长时间(h)101112131415sumFP空质粒组x0.605 0.608 0.616 0.610 0.608 0.618 0.411 3 593.6810.000 s0.018 0.007 0.010 0.011 0.014 0.011 0.272 重组质粒组x1.015 1.057 1.097 1.103 1.164 1.159 0.658 4 125.7910.000 s0.023 0.017 0.027 0.026 0.056 0.022 0.476 sumx0.809 0.832 0.856 0.856 0.885 0.889 0.534 *8 351.248* 0.000*s0.212 0.231 0.248 0.254 0.289 0.279 0.371 *t42.664 70.736 49.077 51.358 28.815 66.182 4 416.540* P0.000 0.000 0.000 0.000 0.000 0.000 0.000 *
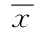
2.5pET30a-Rv2029c促大肠埃希菌生长曲线 以生长时间为横坐标,OD值为纵坐标,绘制出大肠埃希菌生长曲线见图3。在5 h时(此时OD值约为0.45)加入IPTG,空质粒组(pET30a)大肠埃希菌在6 h进入平台期,而重组质粒组(pET30a-Rv2029c)却能继续维持对数期达10 h左右,并且突破大肠埃希菌的生长界限,使大肠埃希菌的OD值达到1.16左右才进入平台期,若以OD值反映大肠埃希菌数量,则重组质粒(pET30a-Rv2029c)能促使大肠埃希菌生长量提高1倍。
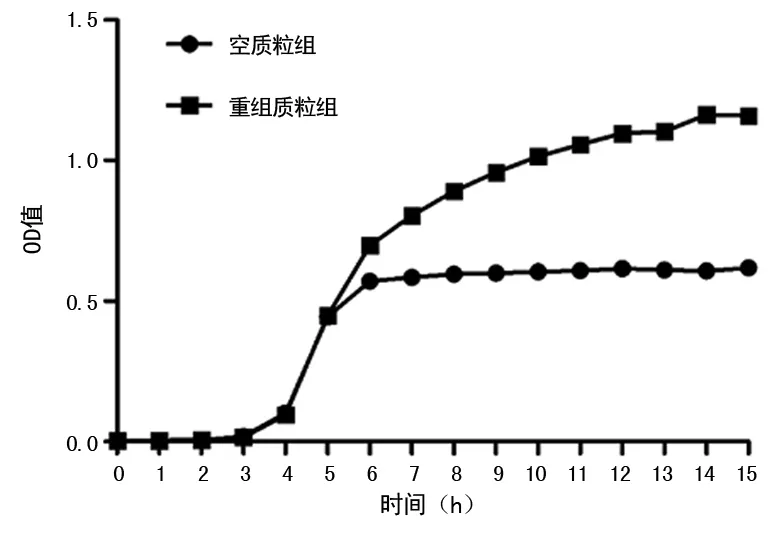
图3 蛋白Rv2029c促大肠埃希菌生长曲线
3 讨 论
本实验发现导入了重组质粒pET30a-Rv2029c的大肠埃希菌与不导入该基因的大肠埃希菌相比,存在着某种机制使大肠埃希菌能够在低氧、营养物质匮乏的环境下继续维持在对数生长期,增强了大肠埃希菌抵抗应激环境的能力,增加了大肠埃希菌的数量。Rv2029c蛋白在Mg2+和ATP的参与下催化6磷酸果糖形成2,6二磷酸果糖,因此在糖代谢中起着重要作用。含有重组质粒pET30a-Rv2029c的大肠埃希菌可能通过Rv2029c蛋白,调节营养物质的供给,提高了对抗低氧环境,增加了利用糖酵解的能力,与不含Rv2029c的大肠埃希菌相比突破了糖代谢和糖利用的上限,促进了大肠埃希菌的生长。
结核分枝杆菌的肉芽肿环境同大肠埃希菌平台期环境相似,都存在营养物质匮乏、低氧、代谢产物增加等对细菌生存不利的因素,依此推断,结核分枝杆菌蛋白Rv2029c可能在肉芽肿环境组织中起到重要的维持细菌存活的作用。结核分枝杆菌在潜伏期为适应肉芽肿的微环境,一方面需要减少自身代谢活动从而进入休眠期,KUMAR等[13]对DosR蛋白中的Rv0079(休眠相关翻译抑制子DATIN) 的研究证实了这一点,发现此蛋白抑制了大肠埃希菌的生长,这是因为该蛋白与核糖体30S亚基结合从而阻断了蛋白合成中的翻译过程,降低了细菌的繁殖速度,使其进入休眠期。另一方面结核分枝杆菌可能需要最大限度利用环境中营养物质、氧气等满足自身生存基本需要,本文研究的结果支持了这一点,生物信息学分析表明蛋白Rv2029c为6-磷酸果糖激酶2,此蛋白属于pfkB糖激酶超家族,是糖代谢的一种重要信号酶,通过影响2,6二磷酸果糖的水平实现对糖酵解通路的调节,增强了结核分枝杆菌在肉芽肿环境中最大程度利用氧气和营养物质的能力,维持结核分枝杆菌生存基本需求。进一步研究该蛋白促进细菌糖酵解的具体机制是下一步研究的方向。
猜你喜欢
中国医学装备(2016年6期)2016-12-01 06:44:31
西南国防医药(2016年7期)2016-12-01 06:01:30
中国中西医结合皮肤性病学杂志(2016年4期)2016-07-18 10:59:50
中华灾害救援医学(2015年7期)2016-01-07 05:45:21
结核与肺部疾病杂志(2015年3期)2015-07-18 11:08:56
家庭医药(2015年11期)2015-05-30 13:37:08
现代检验医学杂志(2015年2期)2015-02-06 02:00:56
中华皮肤科杂志(2014年3期)2014-12-19 12:54:47
中老年健康(2014年7期)2014-05-30 09:01:27
现代检验医学杂志(2014年1期)2014-02-06 01:29:37
