
The first complete organellar genomes of an Antarctic red alga,Pyropia endiviifolia: insights into its genome architecture and phylogenetic position within genus Pyropia (Bangiales,Rhodophyta)*
2018-08-02 02:51XUKuipeng徐奎鹏TANGXianghai唐祥海BIGuiqi毕桂萁CAOMin曹敏WANGLu王璐MAOYunxiang茅云翔
关键词:王璐
XU Kuipeng (徐奎鹏) , TANG Xianghai (唐祥海) , , BI Guiqi (毕桂萁) ,CAO Min (曹敏), WANG Lu (王璐) , MAO Yunxiang (茅云翔) ,
1 Key Laboratory of Marine Genetics and Breeding (Ocean University of China), Ministry of Education, Qingdao 266003, China 2 Laboratory for Marine Biology and Biotechnology, Qingdao National Laboratory for Marine Science and Technology, Qingdao 266003, China
Abstract Pyropia species grow in the intertidal zone and are cold-water adapted. To date, most of the information about the whole plastid and mitochondrial genomes (ptDNA and mtDNA) of this genus is limited to Northern Hemisphere species. Here, we report the sequencing of the ptDNA and mtDNA of the Antarctic red alga Pyropia endiviifolia using the Illumina platform. The plastid genome (195 784 bp, 33.28%GC content) contains 210 protein-coding genes, 37 tRNA genes and 6 rRNA genes. The mitochondrial genome (34 603 bp, 30.5% GC content) contains 26 protein-coding genes, 25 tRNA genes and 2 rRNA genes. Our results suggest that the organellar genomes of Py. endiviifolia have a compact organization.Although the collinearity of these genomes is conserved compared with other Pyropia species, the genome sizes show signi ficant differences, mainly because of the different copy numbers of rDNA operons in the ptDNA and group II introns in the mtDNA. The other Pyropia species have 2–3 distinct intronic ORFs in their cox 1 genes, but Py. endiviifolia has no introns in its cox 1 gene. This has led to a smaller mtDNA than in other Pyropia species. The phylogenetic relationships within Pyropia were examined using concatenated gene sets from most of the available organellar genomes with both the maximum likelihood and Bayesian methods. The analysis revealed a sister taxa affiliation between the Antarctic species Py. endiviifolia and the North American species Py. kanakaensis.
Keyword: Antarctic; Pyropia endiviifolia; plastid and mitochondrial genomes; genome structure; phylogenetic
1 INTRODUCTION
The evolution of plastids and mitochondria by endosymbiosis is a central dogma of modern eukaryotic cell biology. Both plastids and mitochondria possess their own genomes. Plastids are the lightgathering organelles of algae and plants responsible for photosynthesis, whose origin can be traced back to cyanobacteria (Reyes-Prieto et al., 2007). This photosynthetic organelle is commonly believed to have a single origin in the common ancestor of the Archaeplastida, which comprises glaucophytes, red algae (Rhodophyta), and Viridiplantae (Rodríguez-Ezpeleta et al., 2005). Red algae have the most generich and cyanobacteria-like plastid genomes, followed by glaucophytes and green algae. The mitochondrial genome is a remnant of a eubacterial genome, derived speci fically from within the α-Proteobacteria (Gray et al., 2001). Mitochondria play a crucial role in providing cellular energy (Ogihara et al., 2005). During the course of evolution, this endosymbiont has transferred many of its important genes to the nuclear genome(Taanman, 1999).
The Bangiales order of red algae consists of more than 190 species (Guiry and Guiry, 2017), which are distributed worldwide from tropical seas to polar seas.It is divided into at least fifteen genus-level taxa( Bangia, ‘ Bangia’ 1, ‘ Bangia’ 2, ‘ Bangia’ 3,Boreophyllum, Clymene, Dione, Fuscifolium, Lysithea,Minerva, Miuraea, Porphyra, Pseudobangia, Pyropia,and Wildemania) (Sutherland et al., 2011). The coldwater seaweed genus Pyropia includes the most economically important marine crops grown in intertidal habitats, among which Py ropia haitanensis and Py. yezoensis are widely harvested and traded in East Asian countries, such as China, Korea and Japan(Mumford and Miura, 1988). Pyropia endiviifolia (A.Gepp & E. Gepp) H. G. Choi & M. S. Hwang grows on the Antarctic islands and has been recorded on the Antarctic Peninsula, the South Orkney Islands, the South Shetland Islands and South Georgia Island (Wiencke and Clayton, 1998). This species is olive-green in color, which led to its first speci fic name, Monostroma endiviifolium A & E Gepp (Chamberlain, 1963); this was later revised to Pyropia (Sutherland et al., 2011).
Previous studies have revealed that a limited number of available DNA sequences results in relatively little genetic variation, which can present difficulties in phylogenetic resolution or species identi fication (Dutcher and Kapraun, 1994; Niwa et al., 2004; Xie et al., 2010; Sutherland et al., 2011). In recent decades, with the rapid development of nextgeneration DNA sequencing technologies, it has become convenient to assemble complete organelle genomes from total genomic DNA sequences at relatively low cost, especially for Pyropia species,which have a high proportion of organellar DNA relative to nuclear DNA (Wang et al., 2013). Complete organellar genome sequence information is not only important for genetic breeding but also for evolutionary studies. Phylogenomics is a useful tool for providing evolutionary information for species identi fication,taxonomy and phylogenetic analysis (Henry, 2005;Verbruggen et al., 2010; Janouškovec et al., 2013;Yang et al., 2015; Lee et al., 2016). However, most of the available information about Pyropia ptDNAs and mtDNAs is limited to Northern Hemisphere species,and surprisingly little is known about the organellar genomes of Southern Hemisphere species.
Here, we present the complete organellar genomes of the Antarctic species Py. endiviifolia, which were obtained using the Illumina sequencing technology,and examine its genomic features. Through comparative genomics and phylogenomic analyses,we sought to explore the genome structure and reconstruct the phylogenetic relationships among representative species.
2 MATERIAL AND METHOD
2.1 Collection of samples and morphological observations
Fresh thalli of Py. endiviifolia were collected on February 22, 2014 from intertidal transects along a rocky coastline at Fildes Peninsula, King George Island, Antarctica (62°12′S, 58°57′W). Morphological characters including thallus shape, color, texture and reproductive tissues of the specimens were examined and photographed using an Olympus BX51 microscope (OLYMPUS, Tokyo, Japan).
2.2 DNA extraction, sequencing and genome assembly
Total DNA was extracted from 10 g of frozen thallus material according to the CTAB method(Porebski et al., 1997). Puri fied DNA (5 μg) was fragmented and used to construct short-insert PCR-free libraries following the instructions of the Illumina TruseqTMDNA Sample Preparation Kit (Illumina, San Diego, CA, USA) and was sequenced on an Illumina Genome Analyzer. Adapters and low-quality reads(with ambiguous bases, N; length < 100 bp) were removed using the NGS QC Toolkit (Patel and Jain,2012). The pre-processed sequences were first assembled into non-redundant contigs using Edena with default settings (Hernandez et al., 2008). Then,all contigs were mapped to the reference genomes of Py. haitanensis (NC_007932.1 and NC_017751)using the BLAST program (http://blast.ncbi.nlm.nih.gov/) with an e-value of 1e-5 and the order of the aligned contigs was veri fied. Finally, gaps between the contigs were filled by iterative contig extension using the PRICE software (Ruby et al., 2013). To evaluate the quality of the organelle genome sequences,especially the junctions, validation through intensive PCR-based sequencing was carried out on ABI 3730 instrument by randomly designing 20 pairs of primers(Table S1). The PCR sequences and assembled genomes were aligned using MEGA 6.0 to determine the accuracy of the assembly (Tamura et al., 2013).The complete Py. endiviifolia plastid and mitochondria genomes are available for download via GenBank with accession numbers KT716756 and KU356193.

Fig.1 Pyropia endiviifolia
2.3 Genome annotation and analysis
The organellar genomes were annotated using ORF- finder (http://www.ncbi.nlm.nih.gov/projects/gorf/) and aligned via BLASTX and BLASTN searches at the NCBI website (http://blast.ncbi.nlm.nih.gov/). tRNAs were identi fied using the tRNAscan-SE 1.21 web server (http://lowelab.ucsc.edu/tRNAscan-SE/) and rRNAs were identi fied using the RNAmmer 1.2 server (http://www.cbs.dtu.dk/services/RNAmmer/). Genome maps were drawn with OGDraw (Lohse et al., 2007). Multiple genomes were aligned using MAFFT version 5 and visualized using the mVISTA tool (Mayor et al., 2000; Katoh et al.,2005). A structure comparison was generated by Mauve with the ‘Use seed families’ option (Darling et al., 2004).
2.4 Phylogenetic analyses
To elucidate the phylogenetic position of Py.endiviifolia, the concatenated protein-coding amino acid sequences from both the plastid and mitochondrial genomes were used to construct a phylogenetic tree(Table S2). The genome sequences were aligned using the program MAFFT version 5 and were adjusted manually (Katoh et al., 2005). The aligned sequences were trimmed using trimAl with the option‘automated1’ (Capella-Gutiérrez et al., 2009).Maximum likelihood (ML) analysis was conducted using RaxML-8.2.4 (Stamatakis, 2014). The best model and parameter settings were chosen according to the Akaike information criterion by ProtTest 3.0 for ML analysis (Abascal et al., 2005). The ML searches used the cpREV+G+I substitution model for plastid sequences and JTT+G+I substitution model for mitochondrial sequences (-f a, 1 000 bootstrap replicates). Bayesian analyses were carried out using MrBayes3.2 with the best ProtTest model noted above(Huelsenbeck and Ronquist, 2001). Four independent Markov Chain Monte Carlo chains were run simultaneously and sampled every 100 generations for a total of 1 000 000 generations. The first 10% of the trees were discarded as a “burn-in.”
3 RESULT
3.1 Morphological analysis
On the basis of morphology and life history observations (Wang et al., 2008; Guiry, 2015), the specimen was identi fied as Py. endiviifolia(Chamberlain, 1963; Wiencke and Clayton, 1998).This species was very distinctive in the local region because of its position high in the inter-tidal zone, its rough texture compared with other Pyropia species and its dark greenish color, which became blackish on drying (Fig.1a). The gametophyte blades measured5–20 cm in length and 4–15 cm in width and were monostromatic (Fig.1b, c). They attached to the rocks via abundant rhizoidal cells at the base of the thallus(Fig.1d). The almost colorless spermatangia around the margins of the gametophytes were formed by repeated division of vegetative cells (Fig.1e). The red carposporangia were formed by direct transformation of the vegetative cells while the fertilized zygotosporangium divided mitotically (Fig.1f).
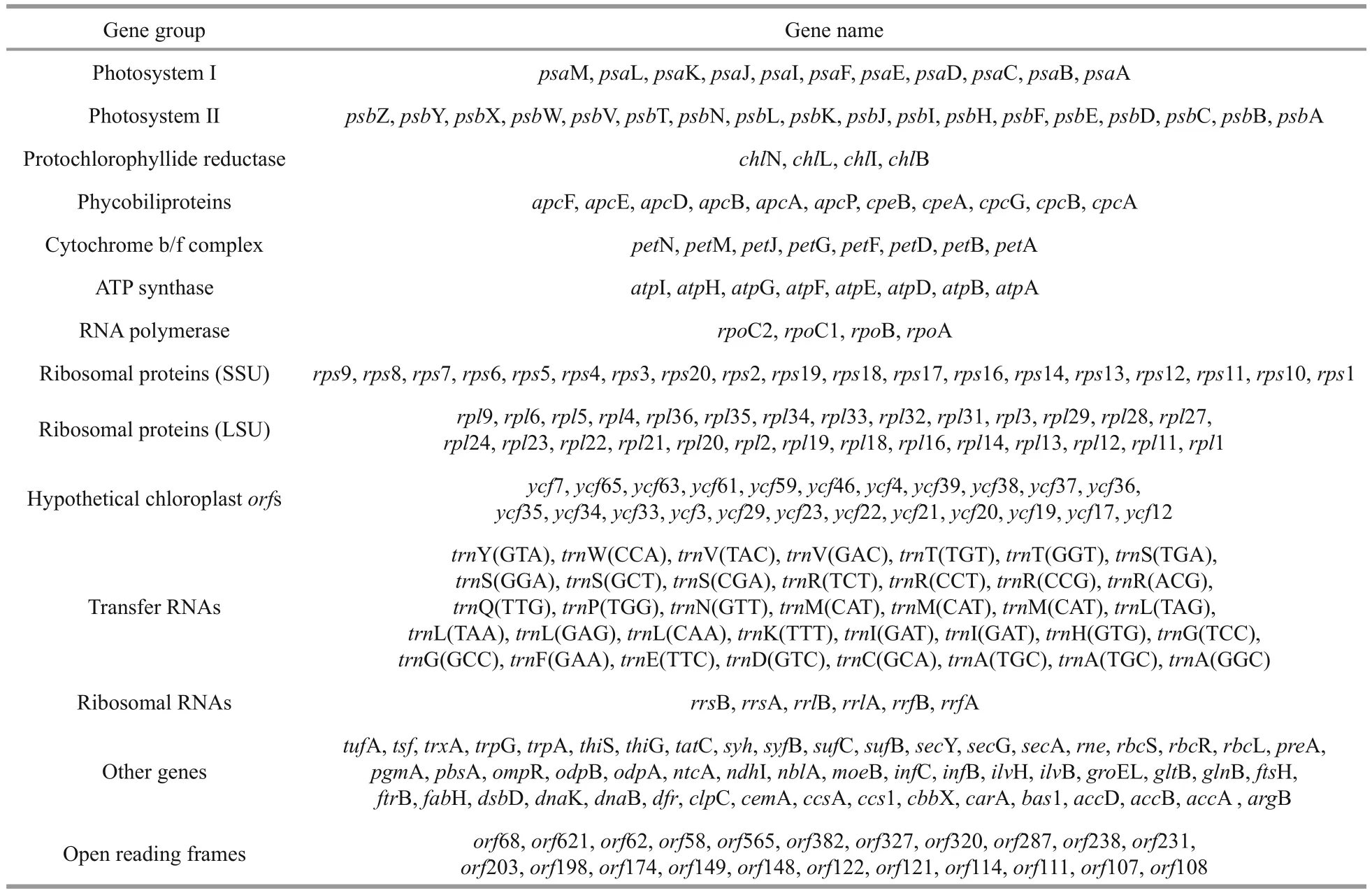
Table 1 Plastid gene content for Pyropia endiviifolia
3.2 Organellar genome features of Py. endiviifolia
The plastid genome of Py. endiviifolia was 195 784 base pairs (bp) long and contained two direct nonidentical repeat (DR) regions encoding 16S, 23S, 5S rRNA and two tRNA genes (trnI, trnA). These two repeats divided the circular molecule into a 150.6-kb large single copy (LSC) region and a 35.6-kb small single copy (SSC) region (Fig.2a). The overall GC content was 33.28%. The plastid genome encoded a total of 253 genes, consisting of 210 protein-coding genes (including 23 hypothetical protein genes ( ycf s)and 22 function-unknown open reading frames( orf s)), 37 tRNA genes and 6 rRNA genes, which comprised 75.83%, 1.44% and 4.58% of the total sequence, respectively (Table 1). Similar to other Bangiales species ( Py. haitanensis, 254 genes; Py.yezoensis, 256 genes; Bangia fuscopurpurea, 250 genes), all the genes in the LSC and SSC regions were single copy without introns, and 14 genes overlapped( psb C–psb D, atp D–atp F, ycf 24–ycf 16, rps 19–rpl 2,rpl 23–rpl 4, car A–orf 238 and rpl 24–rpl 14).
The mitochondrial genome of Py. endiviifolia contained 53 genes, was 34 603 bp in length and had 30.46% GC content (Fig.2b). The genome contained 2 ribosomal RNA genes (1 large subunit and 1 small subunit), 25 transfer RNAs, 3 orf s, 2 secY-independent transporter proteins ( ymf s), 4 ribosomal proteins, and 17 genes related to electron transport and oxidative phosphorylation (Table 2). The protein-coding, tRNA and rRNA genes comprised 58.41%, 5.44% and 18.65% of the whole sequence, respectively.
3.3 Genome conservation and dissimilarities
Multiple alignment of 10 plastid sequences of Bangiales was conducted to further understand the structure and sequence similarity of the Py. endiviifolia plastid genome. Using Py. yezoensis as a reference,the sequence identity alignment results were plotted(Fig.3a). The results revealed high similarity across the Pyropia ptDNAs. The majority of variationsresulted from small insertions or deletions in intergenic regions. As expected, the rDNA regions were more conserved than the single-copy regions,and the coding regions were more conserved than the intergenic regions. Pyropia showed some differences when compared with other genera of Bangiales. For example, the intergenic regions between the pet G-rps 14 genes were longer in the Pyropia ptDNAs.Notably, the similarity of orf 621 was very low between Pyropia and other groups. Collinearity analysis showed that the architecture of the ptDNAs was highly conserved without any large rearrangements, despite their evolutionary distance(Fig.4a). The only apparent distinction was a single
copy rDNA region in the species Py. perforate and Wildemania schizophylla. By contrast, Py. endiviifolia,Py. yezoensis, Py. haitanensis, Porphyra and Bangia possessed two direct non-identical rDNA repeats ( Py.fucicola and Py. kanakaensis had partial genomes).

Table 2 Mitochondrial gene content for Pyropia endiviifolia
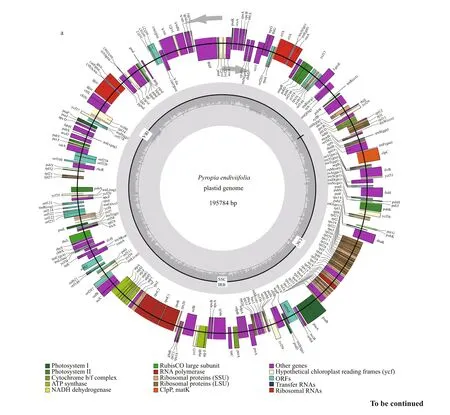
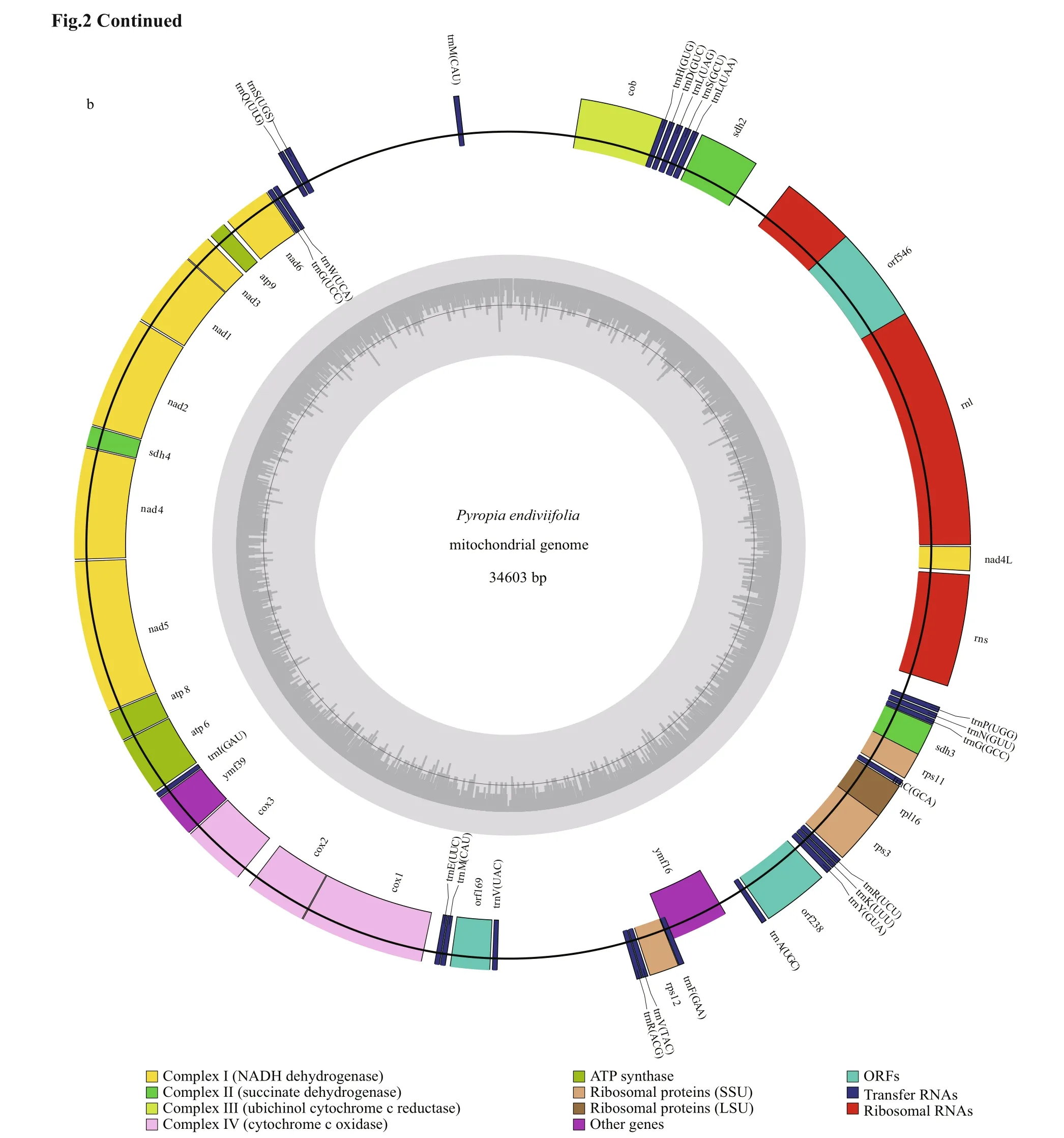
Fig.2 Genome maps of the Pyropia endiviifolia plastid (a) and mitochondrion (b)


Fig.3 Visualization of the alignment of Bangiales organelle genomes sequences

Fig.4 Local collinear block (LCB) analysis of 10 plastid genomes (a) and 12 mitochondrial genomes (b)

Table 3 General characteristics of Bangiales plastid and mitochondrial genomes
Collinearity analysis of 12 mtDNAs showed that most sequence blocks were conserved co-linearly, but the genome contents and lengths were signi ficantly different (Fig.3b; Fig.4b). The observed structural differences mainly arose from the number and organization of mitochondrial group II introns in the large subunit ribosomal RNA ( rnl) gene and the cox 1 gene (Table 3). The mitochondrial genomes of Bangiales possessed different numbers of introns and intronic ORFs ( orf 111, orf 543, orf 544, orf 546,orf 550) in the rnl gene. The mtDNA of Py. endiviifolia had only one intron, which contained one intronic ORF ( orf 546), in the rnl gene. The other genomes of Pyropia contained at least two introns with 0–3 different intronic ORFs. The Pyropia species had 2–3 distinct introns and intronic ORFs ( orf 693, orf 729,orf 789, orf 813) in the cox 1 gene, except Py.endiviifolia, whose cox 1 gene had no introns. An absence of introns in the cox 1 gene was also observed in the Porphyra and Wildemania mtDNAs. The low number of introns resulted in Py. endiviifolia having the smallest mtDNA within Pyropia. B. fuscopurpurea had the most intronic ORFs in the cox 1 gene ( orf 652,orf 693, orf 780 and orf 813).
3.4 Phylogenetic analyses
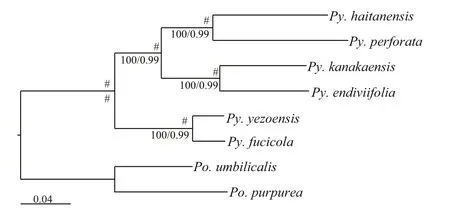
Fig.5 Phylogenetic relationships within the Bangiales clade
Trees were constructed using a dataset of 160 amino acid sequences of ptDNAs and 22 amino acid sequences of mtDNAs selected from eight representative species to examine the evolutionary position of Py. endiviifolia, and all of the nodes were inferred with strong support by the ML and BI methods (Fig.5). Within Pyropia, the close relationship between the Antarctic species Py. endiviifolia and the North American species Py. kanakaensis was con firmed. The species Py. haitanensis and Py.perforate also formed a separate clade. These two clades clustered together with a sister relationship.The remaining species Py. yezoensis and Py. fucicola grouped together at the base of the Pyropia group.

Fig.6 Maximumlikelihoodrootedtree for rbcLsequences of Pyropia
To verify the phylogenetic relationships of this group, the rbc L genes from the Py. endiviifolia plastid genome and 81 species of Pyropia (Table S3)downloaded from GenBank were used for phylogenetic tree reconstruction (Fig.6). The overall topologies were consistent with the trees constructed using whole organellar genomes. The inconsistencies resulted from the low number of available organellar genomes of Pyropia. The phylogenetic tree topology demonstrated that Py. endiviifolia formed a wellsupported clade together with the unidenti fied Pyropia sp. Antar 68 from Admiralty Bay, King George Island,South Shetlands Archipelago, Antarctica. The sequence similarity between Py. endiviifolia and Pyropia sp. Antar 68 was 100%, which indicated that they might be the same species.
4 DISCUSSION
The first plastid and mitochondrial genomes of the Antarctic red algae Py. endiviifolia were determined in this study. The organellar genomes of Py.endiviifolia have large protein-coding gene repertoires and a compact genome organization. Comparative genomic analysis revealed highly conserved collinearity across the whole organellar genomes. The differences in size among mitochondrial genomes were related to the number and organization of mitochondrial group II introns of the large subunit of the ribosomal RNA gene and the cox 1 gene. Typically,eukaryotes possess inserted sequences termed group II introns, but these sequences are only observed in organellar genomes (Michel et al., 1982). A previous study indicated that horizontal transfers have taken place from the mitochondrial genomes of diatoms to the alga Chattonella (Kamikawa et al., 2009). Pyropia endiviifolia had no introns in its cox 1 gene, which was unique among Pyropia species. The number of introns in the rnl gene was also lower than in other Pyropia species. This lack of introns could lead to a convergent and stabilized mtDNA structure. It could also be used as a basis for designing molecular markers for species identi fication. The structure and number of rnl and cox 1 introns in Py. endiviifolia implies a speci fic evolutionary mechanism in this Antarctic species.
We identi fied two direct non-identical repeats in the Py. endiviifolia plastid genomes. By comparison,there was only one copy in Py. perforate and W.schizophylla, a phenomenon that has also been found in some Florideophyte species ( Calliarthron tuberculosum and Chondrus crispus). Typically, most plastid genomes possess two large inverted repeats containing the rRNA genes. However, with the number of sequenced genomes increasing, more and more variations have been found (Hagopian et al.,2004). Analysis of the rDNA operons in Guillardia and Porphyra suggests that the directly repeated rDNA genes of the ancestral Rhodophyte were transformed into inverted repeats in Guillardia(Douglas, 1998). Lee et al. detected three minor structural types (R1-, R2-, and R3-type) in the Florideophyceae group, which were explained by recombination events of the duplicated rDNA operons(Lee et al., 2016). The two rDNA operons have been only partially retained or one copy has been completely lost in some red algae species. This process could lead to structural stabilization of the plastid genomes. The ancestral R1-type rDNA operon was retained in Py.endiviifolia, which implies slow evolution of the ptDNA structure.
Studies have shown that multigene phylogenies can elucidate phylogenetic relationships more exactly when the different evolutionary rates of the genes are considered (Yoon et al., 2006; Verbruggen et al.,2010). We utilized a set of ptDNA and mtDNA genes to explore the phylogenetic relationships of Pyropia.In this study, the Antarctic species Py. endiviifolia and the North American species Py. kanakaensis grouped together with high support in the phylogenetic analysis, rather than all the of Northern Hemisphere species clustering together first. Despite their geographical isolation, all members of this group are cold-water adapted (Brodie and Irvine, 2003), which has led to almost identical environmental selection pressure. The fixation rate in genome evolution depends on the purifying selection of the environment(Buschiazzo et al., 2012). This result implies that the selection pressure the Antarctic species has experienced was more similar to that of Py.kanakaensis than other species. However,inconsistencies in phylogenetic analysis can occur when there is sparse taxon sampling (Zhao et al.,2016). The current evidence is insufficient to interpret the origin and evolution of Py. endiviifolia, because it is the only Southern Hemisphere species with complete organellar genomes available. Therefore,more taxon information needs to be obtained and further studies combining the nuclear, plastid and mitochondrial genomes need to be performed to better understand the relationship of this algal group.
5 DATA AVAILABILITY STATEMENT
The authors declare that all data supporting the findings of this study are available within the methods and appendix sections.
猜你喜欢
Chinese Physics B(2022年6期)2022-06-29
快乐学习报·教师周刊(2022年13期)2022-05-16
Chinese Physics B(2022年1期)2022-01-23
交通科技与管理(2021年13期)2021-09-10
健康之家(2021年19期)2021-05-23
应用数学(2020年3期)2020-07-28
声屏世界(2020年3期)2020-05-09
伙伴(2019年8期)2019-09-24
 Journal of Oceanology and Limnology2018年4期
Journal of Oceanology and Limnology2018年4期
- Journal of Oceanology and Limnology的其它文章
- Editorial Statement
- Recent insights into physiological responses to nutrients by the cylindrospermopsin producing cyanobacterium,Cylindrospermopsis raciborskii*
- Response of Microcystis aeruginosa FACHB-905 to different nutrient ratios and changes in phosphorus chemistry*
- In fluence of light availability on the speci fic density, size and sinking loss of Anabaena flos- aquae and Scenedesmus obliquus*
- Application of first order rate kinetics to explain changes in bloom toxicity—the importance of understanding cell toxin quotas*
- Regime shift in Lake Dianchi (China) during the last 50 years*